
Mutation
|
|
Den här artikeln behöver källhänvisningar för att kunna verifieras. (2017-05) Åtgärda genom att lägga till pålitliga källor (gärna som fotnoter). Uppgifter utan källhänvisning kan ifrågasättas och tas bort utan att det behöver diskuteras på diskussionssidan. |
| Denna artikel ingår i en artikelserie |
| Genetik |
|---|
 |
| Nyckelämnen |
| Kromosom · DNA · RNA · Genom · Ärftlighet · Mutation · Nukleotid |
| Relaterade ämnen |
| Evolution · Populationsgenetik · Kvantitativ genetik · Molekylär genetik |
| Forskning och tillämpningar |
| Genetiker · DNA-sekvensering · Genteknik |
|
|

Mutationer är förändringar i cellers genetiska material (oftast DNA, ibland RNA). Mutationer har flera olika orsaker, till exempel slumpmässiga kopieringsfel under celldelningen, strålning eller inverkan av vissa kemiska ämnen och virus. Mutationer ingår också som ett naturligt och nödvändigt led i vissa processer i celler. För flercelliga organismer kan det vara intressant att skilja mellan mutationer i de celler som ger upphov till nya individer (könsceller=gameter) och därmed sprids till efterkommande generationer, och mutationer i andra celler (somatiska celler) som bara berör den aktuella individen. De kan senare ge upphov till cancer. Somatiska mutationer är dock inte bara av ondo. Hos immunförsvaret genomgår B-celler somatisk mutation under en process som kallas för affinitetsmognad. Detta skapar en immunologisk anpassning och ökar dessa cellers förmåga att binda patogener.
Den sortens mutation som berör könsceller är en av förutsättningarna för evolution. Den stora majoriteten av mutationer är negativa eller har ingen större effekt på överlevnad eller andra framgångsfaktorer, och endast ett litet antal är fördelaktiga för avkomman. Genom det naturliga urvalet kommer fördelaktiga mutationer att öka i frekvens på grund av att de gör sina bärare mer framgångsrika i att producera avkomma, medan de som är negativa av motsvarande anledning mer eller mindre snabbt kommer att försvinna ur populationen. De mer neutrala mutationerna kommer också efter hand att ansamlas i populationen (i en stabil population fördubblas antalet avkommor för en genomsnittlig individ för varje generation) och göra att det finns en spridning på olika egenskaper. Denna spridning och ansamling av neutrala mutationer är en viktig ingrediens i den moderna evolutionsteorin.
Beskrivning
[redigera | redigera wikitext]Delar av ett visst protein fungerar som enheter, varje med en specifik och självständig funktion som kan kombineras för att producera gener som kodar för nya proteiner med nya egenskaper.[1] Till exempel använder det mänskliga ögat fyra gener för att skapa strukturer som känner av ljus: tre för färgseende, tappcellerna, och en för mörkerseende, stavcellerna; alla fyra uppstod från en enda gen från våra förfäder.[2] En annan fördel av att duplicera en gen (eller ett helt genom) är att detta ökar chansen för att den fungerande uppsättningen finns kvar; det tillåter en gen i paret att förvärva en ny funktion medan den andra kopian utför ursprungsfunktionen.[3][4] Andra typer av mutationer skapar emellanåt nya gener från tidigare icke-kodande DNA.[5][6]
Förändringar i kromosomantalet kan innefatta även större mutationer, där segment i DNA-kedjan, inom kromosomen, går sönder och omordnas. Till exempel, i hominiderna sammansmälte två kromosomer för att producera det andra mänskliga kromosomparet; denna sammansmältning skedde inte i den evolutionära utvecklingslinjen hos de andra aporna, vilka fortfarande behåller dessa särskilda kromosomer.[7] Evolutionsmässigt kan den viktigaste uppgift som sådana kromosomomlagringar har vara att påskynda avvikelsen inom en population till att bli nya arter genom att göra populationerna mindre sannolika att para sig, och därigenom bevara genetiska olikheter mellan populationerna.[8]
Sekvenser av DNA som kan röra sig inom genomet, såsom transposoner, utgör en stor del av det genetiska materialet i växter och djur, och kan ha spelat en stor roll i de evolutionära genomen.[9] Till exempel är mer än en miljon kopior av Alu-sekvenser en del av det mänskliga genomet, dessa sekvenser har lagts till för att utföra funktioner så som att reglera genuttryck.[10] En annan effekt av dessa mobila DNA-sekvenser är att när de rör sig inom ett genom, kan de mutera eller ta bort befintliga gener och därigenom skapa mångfald bland generna.[11] Mutationer som inte är dödliga ackumuleras inom genpoolen och ökar den genetiska variationen.[12] Överflödet av vissa genetiska förändringar inom genpoolen kan reduceras av naturlig urval, medan andra ”gynnsammare” mutationer kan ackumuleras och resultera i adaptiva förändringar.
Till exempel kan en fjäril producera en avkomma med nya mutationer. Majoriteten av dessa mutationer kommer inte att ha någon påverkan; men en mutation kan möjligen ändra färgen på en av fjärilens avkommor, och genom det göra det lättare eller svårare för predatorer att uppmärksamma fjärilen. Om färgförändringen är fördelaktig är chanserna för fjärilens överlevnad och reproduktion lite större och över tid kan fjärilar som bär på mutationen utgöra en större andel av populationen.
Neutrala mutationer är definierade som mutationer vars effekter inte påverkar individens anpassbarhet. Dessa mutationer kan öka i antal över tid på grund av genetisk drift. Man tror att den absoluta majoriteten av mutationer inte har någon speciell effekt på organismens anpassbarhet.[13][14] Repareringsprocesser i DNA har möjligheten att åtgärda de flesta förändringar i genomet innan de blir permanenta mutationer. Många andra organismer har processer för att eliminera i andra fall bestående mutationer på somatiska celler.
Fördelaktiga mutationer kan öka den reproduktiva framgången.[15][16]
Skadliga mutationer
[redigera | redigera wikitext]Förändringar i DNA orsakade av mutationer kan orsaka helt och hållet dysfunktionella proteiner. För att en cell ska fungera korrekt måste tusentals proteiner fungera på rätt plats vid rätt tid. När en mutation ändrar ett protein som har en viktig roll i kroppen, kan det resultera i medicinska komplikationer; trots att proteinet är annorlunda försöker cellen använda det som förut. En komparativ studie av gener mellan olika arter av Drosophila indikerade att ungefär 70 % av mutationerna innebar skadliga effekter och de resterande 30 % antingen neutrala eller svagt gynnsamma effekter.[17] Studier har visat att bara 7 % av genmutationer i icke-kodande DNA i jäst är skadliga medan 12 % i kodande DNA är skadliga. De resterande mutationerna är antingen neutrala eller svagt gynnsamma.[18]
Om en mutation uppstår i en könscell kan det ge upphov till att avkomman bär mutationen i alla celler. Det är det här som sker vid ärftliga sjukdomar. Om det sker en mutation i en gen som reparerar DNA:t i könscellerna kan det innebära att människor har en större risk att drabbas av cancer. Artikeln DNA repair-deficiency disorder listar 34 gametmutationer. Ett exempel är albinism vilken är en mutation som uppstår i OCA1- eller OCA2-genen. Individer med denna typ av sjukdom är mer benägna att drabbas av olika typer av cancer, andra sjukdomar och försämrad syn. Å ena sidan kan en mutation uppstå i en av organismens somatiska celler. Sådana mutationer kommer att vara befintliga i alla efterkommande celler av samma typ i organismen. Vissa mutationer kan leda till att cellen blir malign och därigenom orsaka cancer.[19]
En defekt på DNA:t kan orsaka felaktigheter när DNA:t replikeras. Detta replikationsfel kan föranleda en genmutation som i sin tur skulle kunna orsaka en genetisk avvikelse. DNA-skador repareras av DNA:ts repareringssystem som finns i cellen. Varje cell har ett antal olika sätt för enzymer att hitta samt reparera skadan i DNA:t. Eftersom DNA kan skadas på många olika sätt spelar DNA:ts processer för att reparera skadan en stor roll i att skydda den egna kroppen från sjukdomar. När en DNA-skada väl har utvecklats till en mutation kan mutationen inte repareras. DNA:ts olika sätt att identifiera en skada reagerar bara på ”onormala” strukturer i DNA:t. När en mutation väl uppstår i en gensekvens tolkas det som en normal sekvens i genomet och kan därför heller inte bli reparerad.
Gynnsamma mutationer
[redigera | redigera wikitext]Mutationer som orsakar ändringar i proteinsekvenser kan vara skadliga för en organism men då och då kan effekterna vara positiva i den givna miljön. Om det inträffar kan mutationen göra det möjligt för organismen att motstå särskilda miljöpåfrestningar bättre eller reproducera sig snabbare än organismer som saknar mutationen. Mutationen tenderar i dessa fall att bli vanligare inom populationen, detta genom naturligt urval.
Till exempel ger en radering av 32 specifika baspar i mänskligt CCR5 HIV-resistens hos homozygoter och fördröjer Aids-utveckling i heterozygoter.[20] En möjlig etiologisk förklaring till den relativt höga förekomsten av CCR5-Δ32 i europeiska populationer är att det uppstod resistans gentemot böldpesten i mitten av 1300-talet. Personer med denna mutation hade större sannolikhet att överleva en infektion och därigenom ökade mutationens förekomst i populationen.[21] Den här teorin skulle kunna förklara varför mutationen inte har hittats i södra Afrika, som fortsatt varit oberördt av böldpesten. En nyare teori indikerar att selektionstrycket på CCR5-Δ32-mutationen var orsakad av smittkoppor istället för böldpesten.[22]
Ett exempel på en skadlig mutation är sicklecellanemi, en blodsjukdom som gör att kroppen producerar en ovanlig typ av det syrebärande proteinet hemoglobin i de röda blodkropparna. En tredjedel av ursprungsbefolkningen i subsahariska Afrika bär på genen, eftersom i områden där malaria är vanligt förekommande är det evolutionsmässigt gynnsamt att endast ha en sicklecellgen.[23] De med bara en av de två alleler av sicklecellanemin har högre resistens mot malaria då angreppet av malarians plasmodium stoppas av cellernas skärliknande form som den angriper.
Mutationstyper
[redigera | redigera wikitext]Deletion (borttagning)
[redigera | redigera wikitext]
Vid deletion försvinner en nukleotid eller flera nukleotider från sekvensen. När detta händer i en sekvens som definierar ett protein kan en effekt bli att alla efterföljande aminosyredefinitioner blir omtolkade genom att nukleotiderna grupperas annorlunda (om antalet nukleotider som försvinner inte är en multipel av tre (3, 6, 9, 12, etc) eftersom nukleotiderna kodar för aminosyror i grupper av tre (tre nukleotider pekar tillsammans ut en aminosyra). Man säger därför att deletion är en frameshift-mutation.
Insertion
[redigera | redigera wikitext]En extra nukleotid eller flera extra nukleotider sätts in. Då trefaldiga (3, 6, 9, 12, 15...) nukleotider sätts in sker en missense-mutation. Ifall mutationen inte är av trefaldig typ sker en frameshift-mutation, vilket skadar hela genen "nedströms".
Substitution
[redigera | redigera wikitext]En nukleotid byts ut. Kan vara en tyst mutation, en missense-mutation eller en nonsensmutation.
Strukturella kromosomförändringar
[redigera | redigera wikitext]-
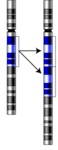 Duplicering
Duplicering -
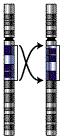 Omvändning
Omvändning -
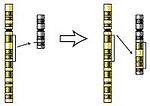 Insertion
Insertion -
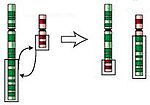 Translokation
Translokation




Mutationer från kromosomavvikelse kan klassificeras efter hur de har påverkat sekvensen av nukleotider. Man talar då om
- Deletion (engelska deletion) - en del av kromosomen förloras
- Duplicering eller duplikation (engelska duplication) - en del repeteras
- Omvändning eller inversion (engelska inversion) - en del vänds
- Insertion (engelska insertion) - en del kopieras och läggs till på en annan plats
- Translokation (engelska translocation) - två delar från olika kromosomer byter plats
Translokation ska inte förväxlas med överkorsning då delar på samma plats i identiska kromosomer byter plats. Överkorsning förekommer alltid vid meios och är en naturlig del av cellens liv.
Numeriska förändringar
[redigera | redigera wikitext]”Sport” i växtförädling
[redigera | redigera wikitext]I växtförädling används stundom termen sport.[24] Det är en spontan mutation i en del av en växt, som sedan förökas vegetativt. Ett exempel är rosenkultivaren Chicago Peace, som är en sport av rosen Peace.[25]
Källor
[redigera | redigera wikitext]- ^ Wang, Minglei; Caetano-Anollés, Gustavo (2009-01). ”The Evolutionary Mechanics of Domain Organization in Proteomes and the Rise of Modularity in the Protein World”. Structure 17 (1): sid. 66–78. doi:. ISSN 0969-2126. http://dx.doi.org/10.1016/j.str.2008.11.008.
- ^ Bowmaker, James K (1998-05). ”Evolution of colour vision in vertebrates”. Eye 12 (3): sid. 541–547. doi:. ISSN 0950-222X. http://dx.doi.org/10.1038/eye.1998.143.
- ^ Dumas, Christian (1999-05). ”The Structure and Reproduction of Corn, 50th Anniversary Edition, T.A. Kisselbach, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1999. US$ 25”. Plant Science 143 (1): sid. 119. doi:. ISSN 0168-9452. http://dx.doi.org/10.1016/s0168-9452(99)00030-8.
- ^ Hurles, Matthew (2004-07-13). ”Gene Duplication: The Genomic Trade in Spare Parts”. PLoS Biology 2 (7): sid. e206. doi:. ISSN 1545-7885. http://dx.doi.org/10.1371/journal.pbio.0020206.
- ^ Liu, Na; Okamura, Katsutomo; Tyler, David M; Phillips, Michael D; Chung, Wei-Jen; Lai, Eric C (2008-08-19). ”The evolution and functional diversification of animal microRNA genes”. Cell Research 18 (10): sid. 985–996. doi:. ISSN 1001-0602. http://dx.doi.org/10.1038/cr.2008.278.
- ^ Siepel, A. (2009-10-01). ”Darwinian alchemy: Human genes from noncoding DNA”. Genome Research 19 (10): sid. 1693–1695. doi:. ISSN 1088-9051. http://dx.doi.org/10.1101/gr.098376.109.
- ^ Zhang, J. (2004-05-01). ”Testing the Chromosomal Speciation Hypothesis for Humans and Chimpanzees”. Genome Research 14 (5): sid. 845–851. doi:. ISSN 1088-9051. http://dx.doi.org/10.1101/gr.1891104.
- ^ Ayala, F. J.; Coluzzi, M. (2005-05-03). ”Chromosome speciation: Humans, Drosophila, and mosquitoes”. Proceedings of the National Academy of Sciences 102 (Supplement 1): sid. 6535–6542. doi:. ISSN 0027-8424. http://dx.doi.org/10.1073/pnas.0501847102.
- ^ Hurst, Gregory D. D.; Werren, John H. (2001-08). ”The role of selfish genetic elements in eukaryotic evolution”. Nature Reviews Genetics 2 (8): sid. 597–606. doi:. ISSN 1471-0056. http://dx.doi.org/10.1038/35084545.
- ^ Häsler, Julien; Strub, Katharina (2006-10-04). ”Alu elements as regulators of gene expression”. Nucleic Acids Research 34 (19): sid. 5491–5497. doi:. ISSN 0305-1048. http://dx.doi.org/10.1093/nar/gkl706.
- ^ Aminetzach, Y. T. (2005-07-29). ”Pesticide Resistance via Transposition-Mediated Adaptive Gene Truncation in Drosophila”. Science 309 (5735): sid. 764–767. doi:. ISSN 0036-8075. http://dx.doi.org/10.1126/science.1112699.
- ^ Eyre-Walker, Adam; Keightley, Peter D. (2007-08). ”The distribution of fitness effects of new mutations”. Nature Reviews Genetics 8 (8): sid. 610–618. doi:. ISSN 1471-0056. http://dx.doi.org/10.1038/nrg2146.
- ^ Ninio, Jacques (1984-05-07). ”The Neutral Theory of Molecular Evolution”. FEBS Letters 170 (1): sid. 210–211. doi:. ISSN 0014-5793. http://dx.doi.org/10.1016/0014-5793(84)81411-8.
- ^ Bohidar, Himadri B. Fundamentals of Polymer Physics and Molecular Biophysics. Cambridge University Press. sid. 192–205. ISBN 9781107415959. http://dx.doi.org/10.1017/cbo9781107415959.013
- ^ Ghiselin, Michael T. (2001-04-05). ”Book Review Dear Mr. Darwin: Letters on the Evolution of Life and Human Nature By Gabriel Dover. 268 pp., illustrated. Berkeley, University of California Press, 2000. $27.50. 0-520-22790-5”. New England Journal of Medicine 344 (14): sid. 1102–1103. doi:. ISSN 0028-4793. http://dx.doi.org/10.1056/nejm200104053441418.
- ^ Tibayrenc, Michel (2003-02). ”Infectious diseases and the Human Genome Diversity Project”. Infection, Genetics and Evolution 2 (3): sid. 165–166. doi:. ISSN 1567-1348. http://dx.doi.org/10.1016/s1567-1348(03)00003-0.
- ^ Sawyer, S. A.; Parsch, J.; Zhang, Z.; Hartl, D. L. (2007-04-04). ”Prevalence of positive selection among nearly neutral amino acid replacements in Drosophila”. Proceedings of the National Academy of Sciences 104 (16): sid. 6504–6510. doi:. ISSN 0027-8424. http://dx.doi.org/10.1073/pnas.0701572104.
- ^ Doniger, Scott W.; Kim, Hyun Seok; Swain, Devjanee; Corcuera, Daniella; Williams, Morgan; Yang, Shiaw-Pyng (2008-08-29). ”A Catalog of Neutral and Deleterious Polymorphism in Yeast”. PLoS Genetics 4 (8): sid. e1000183. doi:. ISSN 1553-7404. http://dx.doi.org/10.1371/journal.pgen.1000183.
- ^ Ionov, Yurij; Peinado, Miguel A.; Malkhosyan, Sergei; Shibata, Darryl; Perucho, Manuel (1993-06). ”Ubiquitous somatic mutations in simple repeated sequences reveal a new mechanism for colonic carcinogenesis”. Nature 363 (6429): sid. 558–561. doi:. ISSN 0028-0836. http://dx.doi.org/10.1038/363558a0.
- ^ Sullivan, A. D.; Wigginton, J.; Kirschner, D. (2001-08-21). ”The coreceptor mutation CCR5 32 influences the dynamics of HIV epidemics and is selected for by HIV”. Proceedings of the National Academy of Sciences 98 (18): sid. 10214–10219. doi:. ISSN 0027-8424. http://dx.doi.org/10.1073/pnas.181325198.
- ^ ”Episode 10: October 2009”. PsycEXTRA Dataset. 22 november 2009. http://dx.doi.org/10.1037/e738612011-001.
- ^ Galvani, A. P.; Slatkin, M. (2003-11-25). ”Evaluating plague and smallpox as historical selective pressures for the CCR5- 32 HIV-resistance allele”. Proceedings of the National Academy of Sciences 100 (25): sid. 15276–15279. doi:. ISSN 0027-8424. http://dx.doi.org/10.1073/pnas.2435085100.
- ^ ”Mental illness: Frequently asked questions (FAQ)”. PsycEXTRA Dataset. 22 november 2008. http://dx.doi.org/10.1037/e429262008-001.
- ^ ”Ordf6rklaring”. Odla.nu. https://www.odla.nu/fragor-svar/ordforklaring. Läst 28 januari 2024.
- ^ J. L (Jack)Harkness (1980). Världens vackraste rosor och hur man odlar dem.. Stockholm: P. A. Norstedt & Söners förlag. sid. 78. ISBN 91-1-804002-0
Se även
[redigera | redigera wikitext]
| ||||||||||||||