
Dinosauria
| Dinosaurios | ||
|---|---|---|
| Rango temporal: 233 Ma - 0 Ma Triásico - Reciente | ||
 Una colección de esqueletos de dinosaurios fósiles. En sentido horario desde la parte superior izquierda: Microraptor gui (un terópodo alado), Apatosaurus louisae (un saurópodo gigante), Edmontosaurus regalis (un ornitópodo con pico de pato), Triceratops horridus (un ceratopsiano con cuernos), Stegosaurus stenops (un estegosauriano plateado), Pinacosaurus grangeri (un anquilosauriano blindado) | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Subfilo: | Vertebrata | |
| Superclase: | Tetrapoda | |
| Clase: | Sauropsida | |
| Subclase: | Diapsida | |
| Infraclase: | Archosauromorpha | |
| (sin rango): | Ornithodira | |
| Superorden: |
Dinosauria Owen, 1842 | |
| Órdenes | ||
Taxones de afinidades inciertas
| ||
Los dinosaurios, del superorden Dinosauria (del griego δεινός deinós ‘terrible’ y σαῦρος sauros ‘lagarto’, ‘lagartos terribles’) son un grupo de saurópsidos[7][8] diápsidos arcosaurios que aparecieron durante el período Triásico con distribución cosmopolita. Aunque el origen exacto y su diversificación temprana es tema de activa investigación,[9] el consenso científico actual sitúa su origen entre 245 y 231 millones de años atrás, entre el Triásico Medio al Superior.[10]
Fueron los vertebrados terrestres dominantes durante 135 millones de años, desde el inicio del Jurásico, hace unos 200 millones de años, a partir de la extinción masiva del Triásico-Jurásico, hasta el final del Cretácico, hace 66 millones de años, cuando la mayoría de los grupos de dinosaurios se extinguieron durante la extinción masiva del Cretácico-Paleógeno que puso fin a la Era Mesozoica. El registro fósil indica que las aves evolucionaron a partir de dinosaurios terópodos durante el período Jurásico y, en consecuencia, muchos taxónomos consideran que las aves forman un subgrupo dentro de los dinosaurios.[11] Algunas aves sobrevivieron a este acontecimiento, y sus descendientes continúan el linaje de los dinosaurios hasta nuestros días.[12] Por lo tanto, los dinosaurios se pueden dividir en dinosaurios aviares, las aves en un sentido estricto, y los dinosaurios no aviares, todos extintos, que son todos dinosaurios distintos de las aves.
Los dinosaurios son un grupo diverso de animales desde el punto de vista taxonómico, morfológico y ecológico. Usando la evidencia fósil, los paleontólogos han identificado cerca de 900 géneros distintos[13] y más de 1000 especies diferentes de dinosaurios no avianos.[14] Los dinosaurios están representados en todos los continentes tanto por especies existentes, las aves, como por restos fósiles.
Algunos son herbívoros, otros carnívoros u omnívoros. Los primeros dinosaurios fueron bípedos, pero muchos grupos incluyeron especies cuadrúpedas, y algunos podían alternar los dos tipos de locomoción. Los cuernos o crestas son comunes a todos los grupos de dinosaurios, y algunos grupos desarrollaron modificaciones esqueléticas como armaduras óseas y espinas. La evidencia sugiere que la puesta de huevos y la construcción de nidos fueron rasgos que compartían todos los dinosaurios. Muchos dinosaurios eran de gran porte, el dinosaurio saurópodo más grande pudo haber alcanzado una longitud de 40 m y 18 m de altura y una masa estimada máxima de 140 t. Sin embargo, la idea de que los dinosaurios no aviares fueron todos gigantescos es un error basado en el sesgo de conservación, ya que los huesos grandes y fuertes tienen más probabilidad de durar hasta que se fosilicen. Muchos dinosaurios eran bastante pequeños, algunos median solamente unos 50 cm de largo.
Aunque la palabra dinosaurio significa ‘lagarto terrible’, el nombre es un poco engañoso, ya que los dinosaurios no son lagartos. En cambio, representan un grupo separado de reptiles que, como muchas formas extintas, no muestran características tradicionalmente vistas como propias de un reptil, como la postura extendida hacia los lados de las extremidades o la ectotermia. Además, muchos animales prehistóricos, incluidos los ictiosaurios, mosasaurios, plesiosaurios, pterosaurios y pelicosaurios, como Dimetrodon, son popularmente concebidos como dinosaurios, pero no están clasificados taxonómicamente como tales. A lo largo de la primera mitad del siglo XX, antes de que las aves fueran reconocidas como dinosaurios, la mayor parte de la comunidad científica creía que los dinosaurios habían sido lentos y de sangre fría. No obstante, la mayoría de las investigaciones realizadas desde la década de 1970 han indicado que todos los dinosaurios eran animales activos con metabolismo elevado y numerosas adaptaciones para la interacción social.[15]
Desde que en el siglo XIX se reconocieron los primeros fósiles de dinosaurio, los esqueletos fósiles montados han sido la principal atracción de muchos museos de todo el mundo, y los dinosaurios se han convertido en parte permanente de la cultura mundial. El gran tamaño de algunos grupos, así como su naturaleza aparentemente monstruosa y fantástica, han asegurado apariciones regulares de dinosaurios en los libros de mayor venta y en películas como Parque Jurásico. El persistente entusiasmo del público por estos animales ha dado lugar a una importante financiación para la investigación científica de los dinosaurios, y los nuevos descubrimientos son dados a conocer regularmente en los medios de comunicación.
Definición
[editar]
Desde el punto de vista de la taxonomía filogenética, los dinosaurios se definen generalmente como el grupo constituido por todos los descendientes del más reciente ancestro común (ACMR) de Triceratops horridus y Passer domesticus, una ave moderna.[16] También se ha propuesto que Dinosauria se defina con respecto al ACMR de Megalosaurus e Iguanodon, porque se trataba de dos de los tres géneros citados por Richard Owen al reconocer la Dinosauria.[17] Ambas definiciones incluyen el mismo grupo de animales: «Dinosauria = Ornithischia + Saurischia», que abarca los terópodos, en su mayoría carnívoros bípedos y aves, anquilosaurianos, cuadrúpedos herbívoros acorazados, estegosaurianos, cuadrúpedos herbívoros con placas en la espalda, ceratopsianos, herbívoros cuadrúpedos con cuernos y volantes, ornitópodos, herbívoros bípedos o cuadrúpedos, incluyendo a los "picos de pato", y sauropodomorfos, en su mayoría grandes cuadrúpedos herbívoros de cuello y cola largos.[18]
En el siglo XXI, las aves se reconocen como las únicas supervivientes del linaje de los dinosaurios terópodos. En la taxonomía tradicional, las aves eran consideradas una clase separada que había evolucionado a partir de los dinosaurios, un superorden distinto. Sin embargo, la mayoría de los paleontólogos contemporáneos relacionados con los dinosaurios rechazan la clasificación de estilo tradicional en favor de la nomenclatura filogenética; este enfoque requiere que, para que un grupo sea natural, todos los descendientes de los miembros del grupo se incluyan también en el grupo. Las aves están, por lo tanto, consideradas dinosaurios, y estos últimos, por consiguiente, no se han extinguido. Las aves se clasifican como pertenecientes al subgrupo Maniraptora, que son coelurosaurios, que a su vez son terópodos, que a su vez son saurisquios, y que a su vez son dinosaurios.[19]
La investigación realizada por Matthew G. Baron, David B. Norman y Paul M. Barrett en 2017 sugirió una revisión radical de la sistemática de los dinosaurios. Análisis filogenético de Baron et al. recuperó los Ornithischia por estar más cerca de los Theropoda que los Sauropodomorpha, en contraposición a la unión tradicional de terópodos con sauropodomorfos. Esto haría que los saurópodos y sus parientes quedaran fuera de los dinosaurios tradicionales, por lo que redefinieron Dinosauria como el último ancestro común de Triceratops horridus, Passer domesticus y Diplodocus carnegii y todos sus descendientes, para garantizar que los saurópodos y sus parientes sigan incluidos como dinosaurios. También resucitaron el clado Ornithoscelida para referirse al grupo que contiene Ornithischia y Theropoda.[20][21][22]
La hipótesis de Ornithoscelida fue cuestionada por un equipo internacional de investigadores en noviembre de 2017, tras haber corregido el conjunto de datos anatómicos original usado por Baron y colaboradores. Esta reelaboración dio como resultado el modelo tradicional, con los grupos Ornithischia y Saurischia recuperados como taxones hermanos. Sin embargo, sus autores reconocieron que este esquema tradicional tiene un apoyo débil y no fue estadísticamente muy diferente de la hipótesis alternativa de Ornithoscelida.[23][24]
Descripción
[editar]Descripción general
[editar]
Los dinosaurios se pueden describir generalmente como arcosaurios con extremidades mantenidas erectas por debajo del cuerpo.[25] Muchos grupos de animales prehistóricos son popularmente concebidos como dinosaurios, como los ictiosaurios, plesiosaurios, mosasaurios, pterosaurios y pelicosaurios, pero no se clasifican científicamente como dinosaurios, y ninguno tenía la postura característica de miembro erecto de los verdaderos dinosaurios.[26] Los pterosaurios están relacionados lejanamente con los dinosaurios, siendo miembros del clado Ornithodira. Los otros grupos mencionados son, al igual que los dinosaurios y los pterosaurios, miembros de Sauropsida, el clado de reptiles y aves, excepto los pelicosaurios, que son sinápsidos. Ninguno de ellos tenía la postura erguida de las extremidades traseras característica de los verdaderos dinosaurios.[26]
Los dinosaurios eran los vertebrados terrestres dominantes del Mesozoico, especialmente en los períodos Jurásico y Cretácico. Otros grupos de animales estaban restringidos en tamaño y nichos. Los mamíferos, por ejemplo, rara vez superaban el tamaño de un gato, y se trataba generalmente de roedores carnívoros del tamaño de una pequeña presa.[27] Se han identificado con certeza más de 900 géneros de dinosaurios no aviares hasta 2018, y el número total de géneros conservados en el registro fósil se ha estimado en alrededor de 1850, casi el 75% de los cuales aún están por descubrir, y se conocen 1124 especies para 2016.[28][29][13] Un estudio anterior predijo que existían alrededor de 3400 géneros de dinosaurios, incluyendo muchos que no se han conservado en el registro fósil.[30] En 2016, el número estimado de especies de dinosaurios que existieron en el Mesozoico era de 1543 a 2468.[31][32] En 2021, se estimó que el número de aves modernas, los dinosaurios aviares, era de 10806 especies.[33]
Algunos eran herbívoros, otros carnívoros, incluyendo granívoros, piscívoros, insectívoros y omnívoros. Aunque los dinosaurios eran ancestralmente bípedos, al igual que todas las aves modernas, algunas especies prehistóricas eran cuadrúpedas, y otras, como Ammosaurus e Iguanodon, podían caminar con la misma facilidad en dos o cuatro patas. Las modificaciones craneales como cuernos y crestas son rasgos comunes de dinosaurios, y algunas especies extinguidas tenían armadura ósea. Aunque conocidos por su gran tamaño, muchos dinosaurios del Mesozoico eran del tamaño de un ser humano o menor, y las aves modernas son en general de pequeño tamaño. Hoy en día se encuentran dinosaurios enterrados en todos los continentes, y los fósiles muestran que habían logrado la distribución global por lo menos durante el período Jurásico temprano.[15] Las aves modernas habitan en la mayoría de los hábitats disponibles, desde terrestres hasta marinos, y existe evidencia de que algunos dinosaurios no aviares, como el Microraptor, podían volar o al menos planear y otros, como los espinosáuridos, tenían hábitos semiacuáticos.[34]
Características anatómicas distintivas
[editar]Si bien los descubrimientos recientes han hecho que sea más difícil presentar una lista universalmente aceptada de sus características distintivas, casi todos los dinosaurios descubiertos hasta ahora comparten ciertas modificaciones en el esqueleto ancestral del arcosaurio, o son claramente descendientes de dinosaurios más antiguos que muestran estas modificaciones. A pesar de que algunos géneros desarrollaron adaptaciones que hicieron aún más pronunciadas las diferencias estructurales, esos rasgos básicos son considerados como típicos del superorden Dinosauria, dichas cualidades comunes a los miembros de un grupo taxonómico reciben el nombre de sinapomorfias de dicho grupo.[35]

Una evaluación detallada de las interrelaciones entre arcosaurios de S. Nesbitt[36] confirmaron o encontraron las siguientes doce sinapomorfias inequívocas, algunas conocidas previamente:
- En el cráneo se presenta una fosa supratemporal, una excavación por delante de la fenestra supratemporal, la principal abertura en la parte superior del cráneo trasero.
- En las vértebras cervicales anteriores, también llamadas frontales, del cuello, detrás del atlas y el axis están presentes las Epipófisis, procesos oblicuos en forma de puntas apuntando hacia atrás en las esquinas superiores traseras.
- En el húmero, hueso del brazo, el vértice de la cresta deltopectoral, una proyección en la que los músculos se unen deltopectorales, situado al nivel o más del 30 % de la longitud del mismo.
- El radio, un hueso del brazo inferior, más corto que el 80 % de la longitud del húmero
- En el fémur, hueso del muslo, el cuarto trocánter, proyección donde el músculo caudofemoralis se une al eje posterior interno, es una brida aguda.
- El cuarto trocánter es asimétrico con el margen distal inferior formando un ángulo más pronunciado con la diáfisis del fémur.
- La faceta articular proximal para la fíbula o peroné del astrágalo y el calcáneo, huesos superiores del tobillo, abarca menos del 30% del ancho transversal del elemento.
- Los exocciptiales, huesos en la parte posterior del cráneo, no se tocan a lo largo de la línea media en el suelo de la cavidad endocraneal, el espacio interior de la caja craneal.
- En la pelvis, las superficies articulares proximales del isquion con el ilion y el pubis están separadas por una gran superficie cóncava, en el lado superior del isquion entre los contactos con el hueso púbico de la cadera y el ilion se encuentra una parte de la articulación abierta.
- Las crestas cnémial, parte sobresaliente de la superficie superior en la tibia, se arquean anterolateralmente, es decir, hacia la parte superior de un lado.
- Cresta orientada proximodistalmente distinta presente en la cara posterior del extremo distal de la tibia.
Nesbitt encontró una serie de potenciales nuevas sinapomorfías, y descontó un número de sinapomorfías sugeridas anteriormente. Algunas de estas también están presentes en los silesáuridos, que Nesbitt recuperó como un grupo hermano de Dinosauria, incluyendo un gran trocánter anterior, metatarsianos II y IV de longitud subequal, reducción de contacto entre el isquion y el pubis, la presencia de una cresta cnemial en la tibia y de un proceso ascendente en el astrágalo, y muchos otros.[16]

j: yugal, p: hueso parietal, po: hueso postorbital, q: hueso cuadrado, qj: hueso quadratojugal, sq: hueso escamosal.
Una variedad de otras características del esqueleto son compartidas por los dinosaurios. Sin embargo, debido a que son comunes a otros grupos de arcosaurios o no estaban presentes en todos los dinosaurios ancestrales, estas características no son considerados como sinapomorfías. Por ejemplo, al igual que los diápsidos, los dinosaurios ancestrales tenían dos pares de fenestra temporal, aberturas en el cráneo detrás de los ojos, y como miembros del grupo diápsido Archosauria, tenían aberturas adicionales en el hocico y la mandíbula inferior. Además, varias características que se creía eran sinapomorfías ahora se sabe que han aparecido antes de los dinosaurios, o estaban ausentes en los primeros dinosaurios y evolucionaron de forma independiente por diferentes grupos de dinosaurios.[37] Estas incluyen una escápula alargada u omóplato, un sacro compuesto por tres o más vértebras fusionadas, en algunos otros arcosaurios se encuentran tres, pero en Herrerasaurus se encuentran solo dos y un acetábulo perforado, o cavidad de la cadera, con un agujero en el centro de su superficie interior, que, por ejemplo en Saturnalia, se presenta cerrado. Otra dificultad de determinar claramente las características de los dinosaurios es que los primeros dinosaurios y otros arcosaurios del Triásico Tardío suelen ser poco conocidos aunque fueron similares en muchos aspectos, estos animales en ocasiones han sido mal identificados en la literatura.[38]

Los dinosaurios se colocaban de pie con sus patas traseras erectas de una manera similar a la mayoría de los mamíferos modernos, pero distinta de la mayoría de los otros reptiles, cuya expansión extremidades a cada lado.[39] también se presentan argumentos y muchas de las imágenes. Esta postura es debida al desarrollo de un rebaje frontal lateral en la pelvis, normalmente una cuenca abierta, y una cabeza distinta correspondiente hacia el interior frontal del fémur.[40] Su posición erecta les permitió a los primeros dinosaurios respirar con facilidad mientras se movían, lo que probablemente les produjo una resistencia extra y niveles de actividad que superaron a los de los "extensos" reptiles.[41] Probablemente las extremidades erectas también ayudaron a apoyar la evolución de gran tamaño mediante la reducción de los esfuerzos de flexión en las extremidades.[42] Algunos arcosaurios no dinosaurianos, incluyendo a los rauisuchianos, también tuvieron las extremidades erectas pero lograron esta característica de «pilar erecto» de la articulación de la cadera, donde el hueso de la pelvis superior, en lugar de tener una proyección de la inserción del fémur desde una toma de la cadera, fue girado para formar una plataforma sobresaliente.[42]
Descubrimiento e investigación
[editar]Historia precientífica
[editar]Los fósiles de dinosaurio son conocidos desde hace milenios, aunque su naturaleza auténtica no fuera entendida. Los chinos, que llaman a los dinosaurios konglong, 恐龍, o ‘dragón terrible’, los consideraban huesos de dragón y los documentaron como tales. Por ejemplo, Hua Yang Guo Zhi, en chino tradicional, 華陽國志, un diccionario geográfico compilado por Chang Qu en chino tradicional, 常璩 durante la Dinastía Jin Occidental, documentaba el descubrimiento de huesos de dragón en Wucheng, en la provincia de Sichuan.[43] Los poblados de China central han excavado huesos de dinosaurios desde hace décadas, pensando que pertenecían a dragones, para hacer medicinas tradicionales.[44] En general, en Europa se creía que los fósiles de dinosaurios eran los restos de gigantes y otras criaturas que murieron en el Diluvio universal.[45]
Investigación temprana sobre dinosaurios
[editar]
Las descripciones académicas de lo que ahora se reconocerían como huesos de dinosaurio aparecieron por primera vez a finales del siglo XVII en Inglaterra. Parte de un hueso, que ahora se sabe que fue el fémur de un Megalosaurus fue el primer dinosaurio en ser descrito formalmente, en 1677, cuando se recuperó una parte de un hueso de una cantera de roca caliza en Cornualles, cerca de Chipping Norton (Inglaterra). Este fragmento óseo fue identificado correctamente como el extremo inferior del fémur de un animal más grande que cualquier ser viviente de tiempos modernos. Identificó correctamente el hueso como la extremidad inferior del fémur de un animal grande, y reconoció que era demasiado grande para pertenecer a cualquier especie conocida. Por lo tanto, concluyó que se trataba del fémur de un humano enorme, tal vez un titán u otro tipo de gigante que aparece en las leyendas.[46][47] Edward Lhuyd, amigo de Isaac Newton, publicó Lithophylacii Britannici ichnographia en 1699, el primer tratamiento científico de lo que ahora sería reconocido como un dinosaurio cuando describió y nombró a un diente de saurópodo, Rutellum impicatum,[48][49] que se había encontrado en Caswell, cerca de Witney, Oxfordshire.[50]
Entre 1815 y 1824, el reverendo William Buckland, primer lector de Geología de la Universidad de Oxford, recolectó más huesos fosilizados de Megalosaurus y se convirtió en la primera persona en describir un dinosaurio no aviar en una revista científica.[51][52] El segundo género de dinosaurio en ser identificado, Iguanodon, fue descubierto en 1822 por Mary Ann Manto, la esposa del geólogo inglés Gideon Mantell. Este reconoció las semejanzas entre sus fósiles y los huesos de las iguanas modernas y publicó sus hallazgos en 1825.[53]
El estudio de estos «grandes lagartos fósiles» pasó a ser de sumo interés para científicos europeos y estadounidenses y en 1842 el paleontólogo inglés Richard Owen creó el término «dinosaurio». Reconoció que los restos que habían sido encontrados hasta ese momento, Iguanodon, Megalosaurus e Hylaeosaurus, compartían un número de rasgos distintivos, de manera que decidió agruparlos como un grupo taxonómico propio. Con el apoyo del príncipe Alberto de Sajonia-Coburgo-Gotha, marido de la Reina Victoria, Owen estableció el Museo de Historia Natural en Kensington Sur, Londres, para exhibir la colección nacional de fósiles de dinosaurios y otros artículos sobre biología y geología.
Etimología
[editar]El taxón Dinosauria fue formalmente nombrado en 1842 por el paleontólogo Richard Owen, que lo utilizó para referirse a la «tribu distinta o sub-orden de los reptiles saurios» que luego fue reconocido en Inglaterra y en todo el mundo.[54] El término deriva de las palabras griegas δεινός, deinós, que significa ‘terrible’, ‘potente’, o ‘gran temor’ y σαῦρος, sauros, que significa ‘lagarto’ o ‘reptil’.[54][55] Aunque el nombre taxonómico ha sido frecuentemente interpretado como una referencia a los dientes, las garras y otras características temibles de los dinosaurios, Owen pretendía simplemente evocar su tamaño y majestuosidad.[56]
Primeros descubrimientos en América del Norte
[editar]En 1858 se descubrió el primer dinosaurio en lo que hoy es el territorio estadounidense en unos pozos de marga en el pequeño pueblo de Haddonfield, Nueva Jersey. Aunque ya se habían encontrado fósiles antes, no se había discernido correctamente su naturaleza. La criatura fue denominada Hadrosaurus foulkii, por el autor del hallazgo, William Parker Foulke. Fue un descubrimiento sumamente importante: el primer esqueleto de dinosaurio encontrado casi completo, el primero había sido descubierto en 1834 en Maidstone (Inglaterra) y demostraba una postura claramente bípeda. Era un descubrimiento revolucionario, ya que hasta entonces, la mayoría de científicos pensaban que los dinosaurios caminaban en cuatro patas, como otros reptiles. Los descubrimientos de Foulke desencadenaron una oleada de «manía de dinosaurios» en Estados Unidos.[57]
Guerra de los Huesos
[editar]


Aquella obsesión por los dinosaurios fue ejemplificada por la gran rivalidad entre Edward Drinker Cope y Othniel Charles Marsh que competían para ver quién descubría más dinosaurios, enfrentamiento que posteriormente fue conocido como la Guerra de los Huesos. La contienda probablemente se originó cuando Marsh indicó públicamente que la reconstrucción de Cope del esqueleto de Elasmosaurus tenía un defecto. Cope sin querer había colocado la cabeza de un plesiosaurio en el lugar donde debería haber sido el extremo caudal del animal. Aquella lucha entre los dos científicos cavadores duró más de 30 años, finalizando en 1897 cuando Cope murió tras haber gastado toda su fortuna en la búsqueda. Marsh ganó la competición principalmente porque estaba mejor financiado debido a su relación con la United States Geological Survey. Desafortunadamente, muchos especímenes valiosos fueron destruidos debido a los toscos métodos de ambos paleontólogos, por ejemplo, sus excavadores muchas veces usaban dinamita para desenterrar huesos, un método que los paleontólogos modernos encontrarían atroz. A pesar de su métodos burdos, las contribuciones de Cope y Marsh a la paleontología fueron enormes. Marsh descubrió 86 nuevas especies de dinosaurios y Cope encontró 56, un total de 142 especies nuevas. La colección de Cope se encuentra a 2016 en el Museo Americano de Historia Natural en Nueva York, mientras que la de Marsh está expuesta en el Museo Peabody de Historia Natural, en la Universidad de Yale.[58]
Primeros descubrimientos en Sudamérica
[editar]Durante la denominada Campaña del Desierto. Junto a los militares marcharon varios científicos de la Academia Nacional de Ciencias de Córdoba. Su misión era hacer un informe sobre la zoología, botánica y geología del nuevo territorio. Durante la ocupación del norte de la Patagonia, las expediciones del comandante Buratowich en 1882, del capitán Rodhe en 1883 y del coronel Romero en 1887 realizaron numerosas contribuciones a la paleontología argentina. Los huesos encontrados durante el mando de Buratowich fueron enviados al entonces presidente Roca, que a su vez se los mostraba al especialista Florentino Ameghino. Los huesos encontrados en la expedición de Rodhe se los enviaron directamente a Ameghino, y los recuperados en la de Romero, se llevaron al Museo de La Plata, donde el propio Ameghino era el subdirector. Aunque Ameghino estaba más familiarizado con el estudio de mamíferos cenozoicos, pudo identificar como restos de dinosaurios los huesos que encontró la expedición de Buratowich en Neuquén. La noticia de este espectacular hallazgo, el primero en toda Sudamérica, lo publicó Ameghino en una carta en el diario La Nación el 23 de marzo de 1883. Desde ese momento, fueron numerosos los hallazgos de huesos de dinosaurios en diversas zonas de la Patagonia y que serían después la base de las primeras publicaciones científicas. Cabe destacar el estudio que realizó el paleontólogo inglés Richard Lydekker quien, en 1893, propone formalmente las primeras especies de dinosaurios de América del Sur. A comienzos del siglo XX se contaba con al menos siete especies de dinosaurios reconocidos.[59]
Primeros descubrimientos en la Antártida
[editar]Desde 1897, la búsqueda de fósiles de dinosaurios se ha extendido a todos los continentes, incluyendo la Antártida. El primer dinosaurio antártico que fue descubierto, el anquilosáurido Antarctopelta Oliveroi, fue encontrado en la isla de Ross en 1986, aunque no fue hasta 1994 que una especie antártica, el terópodo Cryolophosaurus ellioti, fue descrito y nombrado formalmente en una revista científica.
Primeros descubrimientos en Asia
[editar]Hasta comienzos de la década de 1920 y el inicio de las expediciones del Museo Americano de Historia Natural, no se sabía casi nada de los restos fósiles de esta región. Ellas revelaron la existencia en Mongolia de ricos yacimientos de dinosaurios, explorados después por las expediciones rusas y polacas. Los hallazgos procedentes de esta área, en especial los del Desierto de Gobi correspondientes al Cretácico superior, han resultado tan interesantes y apasionantes como los de América del Norte a principios de siglo. Entre los especímenes, cabe señalar Tarbosaurus, Saurolophus, varios anquilosaurianos, paquicefalosaurianos y gran cantidad de pequeños terópodos y grandes saurópodos. En China tras los descubrimientos casuales de Mandschurosaurus en el nordeste del país, a comienzos del siglo XX, vinieron las excavaciones sistemáticas de paleontólogos chinos, lideradas por C. C. Young en las décadas de 1930 y 1940, así como el hallazgo de Lufengosaurus del Jurásico inferior. Les siguieron los trabajos en afloramientos del Cretácico y Jurásico que han producido algunos especímenes bastante espectaculares, sobre todo en la provincia de Sichuán. Desde mediados de la década de 1990 se empezaron a recuperar en la región de Yixian en Liaoning fósiles de varios dinosaurios del Cretácico con una cubierta de plumas en sus cuerpos
Primeros descubrimientos en África
[editar]Entre 1909 y 1912, Werner Janensch realizó para el Museo de Historia Natural de Berlín distintas campañas de excavación en lo que hoy es Tanzania, entonces África Oriental Alemana. Las excavaciones en el Jurásico de Tendaguru reportaron al museo cientos de toneladas de restos fósiles, en su mayoría dinosaurios, entre los que el gigantesco braquiosaurio Giraffatitan es uno de los más conocidos. Los dinosaurios identificados en Tendaguru presentan muchas semejanzas con los que, años antes, Cope y Marsh habían comenzado a describir en la Formación Morrison de los Estados Unidos.
Primeros descubrimientos en Australia
[editar]Los primeros restos fósiles de dinosaurios se conocen de principios del siglo XX. Hasta ahora se han encontrado aquí muy pocos huesos de dinosaurio. Sin embargo, los científicos han descubierto cientos de huellas fascinantes, que podrían anunciar el hallazgo de restos importantes en el futuro.
Primeros descubrimientos de la India
[editar]Los primeros informes sobre dinosaurios procedentes de la India datan de la primera mitad del siglo XIX. Los primeros huesos de dinosaurio en Asia fueron encontrados en la India por un capitán británico de uno de los ejércitos de Compañía Británica de las Indias Orientales en 1828, en Jabalpur, trece años antes de que se acuñara la palabra "dinosaurio". Desde entonces se han encontrado muchos huesos, nidos y huevos en todo el país.
Los lugares de especial investigación de los dinosaurios en 2016 incluyen el sur de Sudamérica (particularmente Argentina) y China. Esta última en particular ha dado muchos ejemplares excepcionales de dinosaurios emplumados debido a la geología única de sus estratos fosilíferos, así como un antiguo clima árido especialmente favorable para la fosilización.
Renacimiento de los dinosaurios
[editar]La Segunda Guerra Mundial provocó una pausa en la investigación paleontológica. Después de la guerra, el centro de atención de la investigación se desvió cada vez más hacia los mamíferos fósiles en lugar de los dinosaurios, a los que se consideraba lentos y de sangre fría.[60][61] Sin embargo, a finales de la década de 1960, el campo de la investigación de los dinosaurios experimentó un aumento en la actividad que continúa.[62] Esto fue provocado, en parte, por el descubrimiento de John Ostrom de Deinonychus, un depredador activo que pudo haber sido de «sangre caliente», endotermia, en contraste marcado con la imagen, aún predominante, de dinosaurios como animales lentos y de «sangre fría», ectotermia. Al mismo tiempo, Robert T. Bakker publicó una serie de estudios que también abogaban por estilos de vida activos en los dinosaurios basados en evidencia anatómica y ecológica, que posteriormente se resumieron en su libro de 1986 The Dinosaur Heresies.
La paleontología de los vertebrados se convirtió en un campo de la ciencia global. Nuevos e importantes descubrimientos de dinosaurios han sido hechos por paleontólogos que trabajan en regiones antes inexploradas, incluyendo India, Sudamérica, Madagascar, la Antártida, y la más considerable, China, los dinosaurios emplumados sorprendentemente bien conservados de China han consolidado aún más la relación entre los dinosaurios y sus descendientes vivientes, las aves modernas. La extensa aplicación de la cladística, que analiza rigurosamente las relaciones entre los organismos biológicos, se ha revelado como extremadamente útil a la hora de clasificar a los dinosaurios. Los análisis cladísticos, junto con otras técnicas modernas, ayudan a compensar el hecho de que el registro fósil está muchas veces incompleto y fragmentario.
Entre terópodos, sauropodomorfos y ornitisquios, el número de géneros nombrados comenzó a aumentar exponencialmente a partir de la década de 1990.[28] A partir de 2008, cada año se nombraban más de 30 nuevas especies de dinosaurios.[63] Al menos los sauropodomorfos experimentaron un aumento adicional en el número de especies nombradas en la década de 2010, con un promedio de 9,3 nuevas especies nombradas cada año entre 2009 y 2020. Como consecuencia, se nombraron más sauropodomorfos entre 1990 y 2020 que en todos los años anteriores combinados.[64] Estas nuevas localidades también condujeron a mejoras en la calidad general de los especímenes, y las nuevas especies reciben cada vez más nombres no de fósiles fragmentados sino de esqueletos más completos, a veces de varios individuos. Mejores especímenes también condujeron a que las nuevas especies fueran invalidadas con menos frecuencia.[63] Las localidades asiáticas han producido los especímenes de terópodos más completos,[65] mientras que las localidades de América del Norte han producido los especímenes de sauropodomorfos más completos.[64]
Antes del renacimiento de los dinosaurios, los dinosaurios se clasificaban principalmente utilizando el sistema tradicional basado en rangos de la taxonomía de Linneo. El renacimiento también estuvo acompañado por la aplicación cada vez más extendida de la cladística, un método de clasificación más objetivo basado en la ascendencia y los rasgos compartidos, que ha demostrado ser tremendamente útil en el estudio de la sistemática y la evolución de los dinosaurios. El análisis cladístico, entre otras técnicas, ayuda a compensar un registro fósil a menudo incompleto y fragmentario.[66][67] Los libros de referencia que resumen el estado de la investigación sobre dinosaurios, como The Dinosauria de David B. Weishampel y sus colegas, hicieron que el conocimiento fuera más accesible[68] y estimuló un mayor interés en la investigación de los dinosaurios. La publicación de la primera y segunda ediciones de The Dinosauria en 1990 y 2004, y de un artículo de revisión de Paul Sereno en 1998, estuvieron acompañadas de un aumento en el número de árboles filogenéticos de dinosaurios publicados.[69]
Preservación molecular y de tejidos blandos
[editar]
Los fósiles de dinosaurios no se limitan a huesos, sino que también incluyen huellas o restos mineralizados de cubiertas de piel, órganos y otros tejidos. De estos, los revestimientos cutáneos basados en proteínas de queratina se conservan más fácilmente debido a su estructura molecular hidrófoba y reticulada.[70] Se conocen fósiles de cubiertas de piel a base de queratina o cubiertas de piel óseas de la mayoría de los grupos principales de dinosaurios. Desde el siglo XIX se han encontrado fósiles de dinosaurios con impresiones de piel escamosa. Samuel Beckles descubrió una extremidad anterior de saurópodo con piel conservada en 1852 que se atribuyó incorrectamente a un cocodrilo, Marsh lo atribuyó correctamente en 1888 y fue objeto de más estudios por parte de Reginald Hooley en 1917.[71] Entre los ornitisquios, en 1884 Jacob Wortman encontró impresiones de piel en el primer espécimen conocido de Edmontosaurus annectens, que fueron en gran parte destruidas durante la excavación del espécimen.[71] Owen y Hooley describieron posteriormente impresiones de la piel de Hypsilophodon e Iguanodon en 1885 y 1917.[72] Desde entonces, las impresiones de escamas se han encontrado con mayor frecuencia entre los hadrosáuridos, donde las impresiones se conocen de casi todo el cuerpo en múltiples especímenes.[73]

.png)
A partir de la década de 1990, importantes descubrimientos de fósiles excepcionalmente conservados en depósitos conocidos como Lagerstätten de conservación contribuyeron a la investigación sobre los tejidos blandos de los dinosaurios.[74][75] Principalmente entre ellas estaban las rocas que produjeron las biotas Jehol, datads del Cretácico temprano y Yanliao, del Jurásico medio a tardío, del noreste de China, de las cuales cientos de especímenes de dinosaurios con impresiones de estructuras parecidas a plumas han sido descritos por Xing Xu y sus colegas.[76][77] En los reptiles y mamíferos vivos, las estructuras celulares que almacenan pigmentos conocidas como melanosomas son parcialmente responsables de producir la coloración[78][79] Se han informado rastros químicos de melanina y melanosomas de formas características en las plumas y escamas de los dinosaurios Jehol y Yanliao, incluidos terópodos y ornitisquios.[80] Esto ha permitido múltiples reconstrucciones de cuerpo completo de la coloración de los dinosaurios, como las de Sinosauropteryx[81] y Psittacosaurus[82] por Jakob Vinther y colegas, y técnicas similares también se han extendido a fósiles de dinosaurios de otras localidades.[78] Sin embargo, algunos investigadores también han sugerido que los melanosomas fosilizados representan restos bacterianos.[83][84] El contenido del estómago en algunos dinosaurios Jehol y Yanliao estrechamente relacionados con las aves también ha proporcionado indicaciones indirectas de la dieta y la anatomía del sistema digestivo.[85][86] Se ha informado más evidencia concreta de la anatomía interna en Scipionyx del Plattenkalk Pietraroja de Italia. Preserva porciones de los intestinos, el colon, el hígado, los músculos y la tráquea.

Al mismo tiempo, una línea de trabajo dirigida por Mary Higby Schweitzer, Jack Horner y sus colegas informaron varios casos de proteínas y tejidos blandos conservados dentro de fósiles de huesos de dinosaurios. Schweitzer y otros habían encontrado varias estructuras mineralizadas que probablemente representaban glóbulos rojos y fibras de colágeno en huesos de tiranosáuridos ya en 1991.[87][88][89] Sin embargo, en 2005, Schweitzer y sus colegas informaron un fémur de Tyrannosaurus que en su interior se conservaba tejido blando y flexible, incluidos vasos sanguíneos, matriz ósea y tejido conectivo,fibras óseas, que habían conservado su estructura microscópica. Este descubrimiento sugirió que los tejidos blandos originales podrían conservarse a lo largo del tiempo geológico,[70] habiéndose propuesto múltiples mecanismos.[90] Más tarde, en 2009, Schweitzer y sus colegas informaron que un fémur de Brachylophosaurus conservaba microestructuras similares, y las técnicas inmunohistoquímicas, basadas en la unión de anticuerpos, demostraron la presencia de proteínas como colágeno, elastina y laminina.[91] Ambos especímenes produjeron secuencias de proteínas de colágeno que eran viables para análisis filogenéticos moleculares, que los agruparon con aves como era de esperar.[91][92] También se ha informado de la extracción de ADN fragmentario para ambos fósiles,[93] junto con un espécimen de Hypacrosaurus.[94] En 2015, Sergio Bertazzo y sus colegas informaron sobre la preservación de fibras de colágeno y glóbulos rojos en ocho especímenes de dinosaurios del Cretácico que no mostraron ningún signo de preservación excepcional, lo que indica que el tejido blando puede preservarse con más frecuencia de lo que se pensaba anteriormente.[95] Se han rechazado las sugerencias de que estas estructuras representan biopelículas bacterianas[96], siendo rechazadas,[97] pero la contaminación cruzada sigue siendo una posibilidad difícil de detectar.[98]
Historia evolutiva
[editar]Orígenes e inicios evolutivos
[editar]

Los dinosaurios divergieron de sus ancestros arcosaurios entre mediados a finales del período Triásico, aproximadamente 20 millones de años después de que la extinción masiva del Pérmico-Triásico acabara con un estimado del 95 % de toda la vida en la Tierra.[99][100][101][102] La datación radiométrica de la formación rocosa que contenía los fósiles del género primitivo de dinosaurio Eoraptor ha arrojado una fecha de 231.4 millones de años, estableciendo su presencia en el registro fósil del Triásico.[103] los paleontólogos creen que Eoraptor se parece al ancestro común de todos los dinosaurios;[104][105] es decir, que de ser así los primeros dinosaurios eran animales depredadores pequeños y bípedos.[106]
Los fósiles de dinosaurios bien conocidos más antiguos conocidos a partir de restos sustanciales datan de la época Carniense del período Triásico y se han encontrado principalmente en las Formaciones Ischigualasto y Santa María de Argentina y Brasil respectivamente, y en la Formación Pebbly Arkose de Zimbabue.[107]
La Formación Ischigualasto, fechada radiométricamente entre 231 y 230 millones de años,[108] ha producido el saurisquio temprano Eoraptor, originalmente considerado un miembro de Herrerasauridae[103] pero ahora considerado como un sauropodomorfo temprano, junto con los herrerasáuridos Herrerasaurus y Sanjuansaurus y los sauropodomorfos Chromogisaurus, Eodromaeus y Panphagia,[109] así como el tentativo ornitisquio temprano Pisanosaurus.[110] El probable parecido de Eoraptor con el ancestro común de todos los dinosaurios sugiere que los primeros dinosaurios habrían sido depredadores pequeños y bípedos.[111][104][106] La Formación Santa María, fechada radiométricamente como más antigua, con 233,23 millones de años,[112] ha producido los herrerasáuridos Gnathovorax y Staurikosaurus, junto con los sauropodomorfos Bagualosaurus, Buriolestes, Guaibasaurus, Macrocollum, Nhandumirim, Pampadromaeus , Saturnalia y Unaysaurus.[109] La Formación Pebbly Arkose, que es de edad incierta pero probablemente era comparable a las otras dos, ha producido el sauropodomorfo Mbiresaurus , junto con un herrerasáurido sin nombre.[107] Restos menos bien conservados de los sauropodomorfos Jaklapallisaurus y Nambalia, junto con el saurisquio temprano Alwalkeria, se conocen en las formacionesb Maleri Superior y Maleri Inferior de la India.[113] La Formación Chañares de Argentina , de edad Carniense, preserva ornitodiranos primitivos parecidos a dinosaurios, como Lagosuchus y Lagerpeton, lo que la convierte en otro sitio importante para comprender la evolución de los dinosaurios. Estos ornitodiranos apoyan el modelo de los primeros dinosaurios como depredadores pequeños y bípedos.[109][114] Los dinosaurios pueden haber aparecido ya en la época anisiana del Triásico, hace aproximadamente 243 millones de años, que es la edad del Nyasasaurus de la Formación Manda de Tanzania. Sin embargo, sus fósiles conocidos son demasiado fragmentarios para identificarlo como un dinosaurio o sólo como un pariente cercano.[115] La referencia de la Formación Manda al Anisiano también es incierta. Independientemente, los dinosaurios existieron junto con los ornitodiranos no dinosaurios durante un período de tiempo, con estimaciones que oscilan entre 5 y 10 millones de años [116] a 21 millones de años.[112]
Cuando los dinosaurios aparecieron, ellos no eran los animales terrestres dominantes. Los hábitats terrestres eran ocupados por varios tipos de arcosauromorfos y terápsidos, como los cinodontes y también estaban los rincosaurios. Sus principales competidores eran los pseudosuquios, como los etosaurios, ornitosúquidos y los rauisuquios, los cuales eran más exitosos que los dinosaurios.[117] Muchos de estos animales desaparecerían a lo largo del Triásico, en alguno de los dos eventos de extinción que hubo. El primero, hace cerca de 215 millones de años, vio la desaparición de una gran variedad de arcosauromorfos basales, incluyendo a los protorosaurios. Esto fue seguido por la extinción masiva del Triásico-Jurásico, hace unos 200 millones de años, en la cual se extinguieron varios otros grupos de arcosaurios primitivos, como los etosaurios, los ornitosúquidos, los fitosaurios y los rauisuquios. Los rincosaurios y los dicinodontes sobrevivieron, al menos en algunas áreas, tan tardíamente como el inicio del Noriense e inicios del Rhaetiense, respectivamente,[118][119] pero la fecha exacta de sus extinciones es incierta. Estas pérdidas dejaron como resultado una fauna terrestre compuesta por crocodilomorfos, dinosaurios, mamíferos, pterosaurios y tortugas.[16] Los primeros linajes de dinosaurios primitivos se diversificaron durante las épocas del Carniense y el Noriense en el Triásico, posiblemente al ir ocupando los nichos de los grupos que se extinguían.[18]
Evolución y paleobiogeografía
[editar]La evolución de los dinosaurios tras el Triásico fue acompañada de los cambios en la vegetación y la posición de los continentes. En el Triásico Superior y el Jurásico Inferior, los continentes estaban conectados a una única masa continental conocida como Pangea, y había virtualmente solo un tipo de fauna de dinosaurios compuesto mayormente por carnívoros celofisoides y herbívoros sauropodomorfos primitivos (prosaurópodos).[120] Las plantas gimnospermas (particularmente coníferas), una fuente potencial de comida, se difundieron en el Triásico Superior. Los sauropodomorfos primitivos no tenían mecanismos sofisticados para procesar la comida en la boca, por lo que debieron emplear otros medios para desmenuzar el alimento a lo largo de su tracto digestivo.[121] La homogeneidad general de la fauna de dinosaurios continuó durante el Jurásico Medio y el Jurásico Superior, épocas en las que muchas localidades tenían depredadores tales como los ceratosaurianos, los megalosauroideos y los carnosaurios, mientras que los herbívoros por lo general eran ornitisquios estegosaurios y grandes saurópodos. Ejemplos de estas faunas se hallan en la formación de Morrison en Norteamérica y Tendaguru en Tanzania. Los dinosaurios de China muestran algunas diferencias, con especializados terópodos como los metriacantosáuridos y saurópodos de cuellos inusualmente largos como Mamenchisaurus.[120] Los anquilosaurios y los ornitópodos se estaban volviendo más comunes, pero los prosaurópodos se habían extinguido. Las coníferas y las pteridofitas eran las plantas más comunes. Los saurópodos, como sus antecesores prosaurópodos, no procesaban el alimento oralmente, pero los ornitisquios estaban evolucionando varias formas para tratar la comida en sus bocas, incluyendo posibles estructuras parecidas a mejillas para mantener la comida dentro de la boca y movimientos mandibulares para masticarla.[121] Otro notable evento evolutivo del Jurásico fue la aparición de las aves propiamente dichas, descendientes de los celurosaurios manirraptores.[99]
Durante el Cretácico Inferior y con el sucesivo rompimiento de Pangea, los dinosaurios se empezaron a diferenciar más en cada continente. La primera parte de esta época vio la expansión de los anquilosaurios, los iguanodontianos y los braquiosáuridos a través de Europa, Norteamérica y el norte de África. Estos últimos fueron complementados o reemplazados en África por los grandes terópodos espinosáuridos y carcarodontosáuridos, y los saurópodos rebaquisáuridos y titanosaurios, también hallados en Suramérica. En Asia, los celurosaurios manirraptores como los dromeosáuridos, trodóntidos y ovirraptorosaurios se convirtieron en los terópodos más comunes, mientras que los anquilosáuridos y los ceratopsios primitivos como Psittacosaurus se convirtieron en herbívoros dominantes. Mientras tanto, Australia fue hogar de una fauna de anquilosaurios basales, "hipsilofodóntidos" e iguanodontianos.[120] Los estegosaurios parecen haberse extinto en algún punto entre el final del Cretácico Inferior y el inicio del Cretácico Superior. Un cambio importante que se produjo durante el Cretácico Inferior, y que se amplificaría en el Cretácico Superior, fue la evolución de las plantas con flor. Al mismo tiempo, varios grupos de dinosaurios herbívoros desarrollaron formas más sofisticadas para procesar la comida en la boca. Los ceratopsios emplearon un método para trozar la comida con dientes apilados en baterías dentales, mientras que los iguanodontianos refinaron su método para triturar los alimentos con baterías dentales, que serían llevadas al extremo por los hadrosáuridos.[121] Algunos saurópodos también desarrollaron dichas baterías, de los cuales el mejor ejemplo es el rebbaquisáurido Nigersaurus.[122]
Hubo tres tipos principales de faunas de dinosaurios en el Cretácico Superior. En los continentes septentrionales de Norteamérica y Asia, los principales terópodos eran los tiranosáuridos y varios tipos de terópodos manirraptores pequeños, mientras que los herbívoros eran predominantemente ornitisquios como los hadrosáuridos, ceratopsios, anquilosáuridos y paquicefalosaurios. En los continentes del sur que resultaron del rompimiento de Gondwana, los terópodos más comunes eran los abelisáuridos, y los saurópodos titanosaurios eran los herbívoros más comunes. Finalmente, en Europa, la fauna se componía de dromeosáuridos, iguanodontianos rabdodóntidos, anquilossaurios nodosáuridos, y los saurópodos titanosaurios eran predominantes.[120] Las plantas con flor se irradiaron con mucho éxito,[121] y los primeros tipos de pastos aparecieron al final del Cretácico.[123] Los hadrosáuridos y los ceratopsios se volvieron sumamente diversos en Norteamérica y el oriente de Asia. Los terópodos también desarrollaron formas omnívoras y herbívoras, destacando entre estas a los tericinosaurios y los ornitomimosaurios.[121]
La extinción masiva del Cretácico-Terciario, la cual ocurrió hace aproximadamente 66 millones de años al final del Cretácico, causó la extinción de todos los grupos de dinosaurios exceptuando a las aves neornites (o quizás casi todos, véase sobre esto más abajo). Algunos otros grupos de diápsidos, como los crocodilianos, los sebecosuquios, las tortugas, los lagartos, las serpientes, los esfenodontes y los coristoderos, también sobrevivieron al evento de extinción.[124]
Los linajes sobrevivientes de aves neornites, incluyendo a los ancestros de las modernas ratites, los patos y gallinas, y una gran variedad de aves acuáticas, se diversificaron rápidamente al principio del período Paleógeno, llenando los nichos ecológicos que quedaron vacantes tras la extinción de los grupos de dinosaurios mesozoicos como las aves arborícolas enantiornites, las aves acuáticas hesperornitiformes, e incluso los grandes terópodos terrestres (que sería el caso de Gastornis, los dromornítidos, y las «aves del terror»). Sin embargo, los mamíferos también evolucionaron con rapidez en esa época, y superaron a las aves neornites en la dominación de la mayoría de los nichos terrestres.[125]
Clasificación
[editar]Los dinosaurios (incluyendo las aves) son arcosaurios, como los cocodrilos modernos. Los cráneos diápsidos de los arcosaurios tienen dos agujeros localizados donde se atan los músculos de la mandíbula, llamado fenestra temporal. La mayor parte de reptiles (incluyendo a las aves) son diápsidos. Los mamíferos, con solo una fenestra temporal, son llamados sinápsidos; y las tortugas, sin fenestra temporal, son anápsidos. Anatómicamente, los dinosaurios comparten muchas otras características de arcosaurio, incluidos los dientes que nacen de alvéolos más bien que como las extensiones directas de las mandíbulas. Dentro del grupo Archosauria, los dinosaurios son diferenciados más perceptiblemente por su paso. Las patas de los dinosaurios se extienden directamente bajo el cuerpo y son rectas, mientras que las patas de lagartos y cocodrilos se extienden hacia fuera, sobresaliendo.
Muchas otras clases de reptiles vivieron en la era Mesozoica, en convivencia con los dinosaurios. Algunos de estos son comúnmente, pero incorrectamente, considerados como dinosaurios, incluyendo a los plesiosaurios (que no están relacionados estrechamente con los dinosaurios) y los pterosaurios, que se desarrollaron separadamente de antepasados reptiles de finales del Triásico.
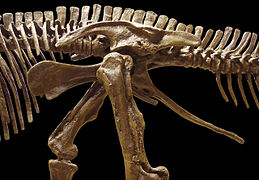
Los dinosaurios están divididos en dos órdenes, Saurischia y Ornithischia, los cuales se basan en la estructura de la cadera. Los saurisquios (del griego para Pelvis de lagarto) son dinosaurios que al principio conservaron la estructura de la cadera de sus antepasados. Ellos incluyen todo los terópodos (carnívoros bípedos) y saurópodos (herbívoros de cuello largo). Los ornitisquios (del griego para Pelvis de ave) son el otro orden de dinosaurios, la mayoría de estos eran cuadrúpedos herbívoros.
-
 Estructura pélvica en Saurischia (lateral izquierdo).
Estructura pélvica en Saurischia (lateral izquierdo). -
 Pelvis saurisquia de Tyrannosaurus (lateral izquierdo).
Pelvis saurisquia de Tyrannosaurus (lateral izquierdo). -
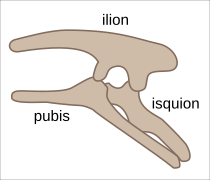 Estructura pélvica en Ornithischia (lateral izquierdo).
Estructura pélvica en Ornithischia (lateral izquierdo). -
 Pelvis ornitisquia de Edmontosaurus (lateral izquierdo).
Pelvis ornitisquia de Edmontosaurus (lateral izquierdo).




Taxonomía
[editar]La siguiente es una clasificación simplificada de los grupos de dinosaurios, en función de sus relaciones evolutivas, y organizados basándose en la lista de especies de dinosaurios mesozoicos facilitados por Holtz (2008).[12] En clasificación de los dinosaurios se puede encontrar una versión más detallada. La cruz (†) se utiliza para significar grupos que no tiene miembros vivientes.
- Dinosauria
- Saurischia («cadera de lagarto»; incluye a Theropoda y Sauropodomorpha)

- †Herrerasauria (primeros carnívoros bípedos)
- †Coelophysoidea (pequeños, primeros terópodos; incluye a Coelophysis y parientes cercanos)
- †Dilophosauridae (primeros terópodos crestados y carnívoros)
- †Ceratosauria (cuernos generalmente elaborados, los carnívoros meridionales dominantes del Cretácico)
- Tetanurae («colas rígidas»; incluye a la mayoría de los terópodos)
- †Megalosauroidea (primeros grupos de grandes carnívoros incluyendo a los espinosáuridos semiacuáticos)
- †Carnosauria (Allosaurus y otros parientes cercanos, como Carcharodontosaurus)
- Coelurosauria («lagartos de cola hueca»; terópodos con plumas, con una amplia gama de nichos y tamaños corporales)
- †Compsognathidae (primeros coelurosaurs comunes con extremidades delanteras reducidas)
- †Tyrannosauridae («lagartos tiranos»;Tyrannosaurus rex y otros parientes cercanos; las extremidades anteriores estaban reducidas)
- †Ornithomimosauria («lagartos imitadores de "avestruz"»; mayormente sin dientes; carnívoros a posibles herbívoros)
- †Alvarezsauroidea (pequeños insectívoros con extremidades anteriores reducidas cada uno con una garra aumentada)
- Maniraptora («con manos de ladrón»; tenían brazos y dedos largos y delgados)
- †Therizinosauria (herbívoros bípedos con grandes garras de mano y cabeza pequeña)
- †Oviraptorosauria («lagartos ladrones de huevos»; mayormente sin dientes; su dieta y estilo de vida son inciertos)
- †Archaeopterygidae (pequeños, terópodos alados o aves primitivas)
- †Deinonychosauria («lagartos corredores»; tamaño pequeño a mediano; similares a las aves, con una garra distintiva en el dedo del pie)
- Avialae («alas de ave»; aves modernas y parientes extintos)
- †Scansoriopterygidae (pequeños aviares primitivos con dedos terceros largos)
- †Omnivoropterygidae («alas omnívoras»; primeros grandes aviares de cola corta)
- †Confuciusornithidae (pequeños aviares sin dientes)
- †Enantiornithes («pájaros opuestos»; primitivos aviares voladores arborícolas)
- Euornithes («aves verdaderas»; aves voladores avanzados)
- †Yanornithiformes (aves chinos dentados del Cretácico)
- †Hesperornithes (aves acuáticas especializadas en inmersiones)
- Aves (modernas, aves picudas modernas y sus parientes extintos)

- †Sauropodomorpha («con forma de pata de lagarto»; herbívoros con cabeza pequeña, cuello y cola larga)
- †Guaibasauridae (pequeños, primitivos, sauropodomorfos omnívoros)
- †Plateosauridae («lagartos planos»; primitivos, bípedos estrictamente "prosaurópodos").
- †Riojasauridae (pequeños, sauropodomorfos primitivos).
- †Massospondylidae (pequeños, sauropodomorfos primitivos).
- †Sauropoda («con pata de lagarto»; muy grandes y pesados, usualmente de más de 15 metros (49,2 pies) de largo; cuadrúpedos)
- †Vulcanodontidae (saurópodos primitivos con extremidades como pilares)
- †Eusauropoda («saurópodos verdaderos»).
- †Cetiosauridae («lagarto ballena»).
- †Turiasauria (Grupo europeo de saurópodos del Jurásico y Cretácico)
- †Neosauropoda («nuevos saurópodos»).
- †Diplodocoidea («lagartos de doble haz»; cráneos y colas alargadas; dientes normalmente estrechos y similares a un lápiz)
- †Macronaria (cráneos cuadrados; dientes de cuchara o en forma de lápiz).
- †Brachiosauridae (macronarianos largamente armados de largos cuellos)
- †Titanosauria (diversos; fornidos, con caderas anchas; más comunes en el Cretácico Tardío de los continentes del sur)

- †Ornithischia («cadera de ave»; diversos herbívoros bípedos y cuadrúpedos)
- †Heterodontosauridae («laragos de dientes diferentes»; pequeños ornitópodos basales herbívoros/omnívoros con prominentes similares a dientes caninos)
- †Thyreophora («portador de escudos»; dinosaurios acorazados; en su mayoría cuadrúpedos)
- †Ankylosauria (escudos como armadura primaria; algunos tenían colas estilo garrote)
- †Stegosauria (pinchos y platos como armadura principal)
- †Neornithischia («nuevos ornitisquios»)
- †Ornithopoda («pie de ave»; diversos tamaños; bípedos y cuadrúpedos; evolucionó un método de mascar usando cráneo flexibles y numerosos dientes)
- †Marginocephalia («cabezas con reborde»; caracterizado por un crecimiento craneal)
- †Pachycephalosauria («lagartos de cabeza gruesa»; bípedos con crecimientos en forma de cúpula o nudos en cráneos)
- †Ceratopsia (presencia de cuernos, un hueso rostral, vértebras del cuello fusionadas y dientes con dos raíces)
Biología
[editar]El conocimiento sobre los dinosaurios ha sido obtenido a través de una variedad de registros fósiles, óseos y no óseos, como huesos fosilizados, huellas de pisadas (icnitas), heces (coprolitos), piedras usadas para ayudar en la digestión (gastrolitos), plumas, impresiones de piel, órganos internos y tejido blando.[126][127] Varios campos de estudio contribuyen a nuestro conocimiento paleontológico de los dinosaurios, incluyendo la física (especialmente la biomecánica; masa de los dinosaurios, velocidad y flujo sanguíneo), química, biología y ciencias de la Tierra (de la que la paleontología es una subdisciplina).[128][129] Dos temas de su particular interés y estudio han sido el tamaño de los dinosaurios y su comportamiento.[130]
Tamaño
[editar]
La evidencia actual sugiere que el tamaño promedio de los dinosaurios varió a través de los períodos Triásico, Jurásico Inferior, Jurásico Superior y Cretácico[104] Los dinosaurios terópodos depredadores, que ocupaban muchos de los nichos de carnívoros terrestres durante el Mesozoico, frecuentemente se ubicaban en la categoría de 100 a 1000 kg (kilogramos) cuando son clasificados por su peso estimado en categorías basadas en el orden de magnitud, mientras que los depredadores mamíferos del Holoceno se ubican en la categoría de 10 a 100 kg.[131] La moda estadística de las masas corporales de los dinosaurios mesozoicos se encuentra entre una a diez toneladas métricas[132] Esto contrasta fuertemente con el tamaño de los mamíferos a lo largo del Cenozoico, cuya masa fue estimada por el Museo Nacional de Historia Natural del Instituto Smithsoniano entre los 2 a 5 kg.[133]
No obstante, el significado de «tamaño promedio» no es tan sencillo de precisar. Las observaciones actuales sugieren diferentes valores para cada uno de los períodos geológicos involucrados.[134]
Los [pesos] estimados de dinosaurios medianos oscilan entre 500 kg y 5 toneladas […] Ochenta por ciento de la biomasa de la Formación de Morrison del oeste de los Estados Unidos consistía en Stegosaurus y saurópodos; estos últimos promediaban las 20 toneladas […]. El típico gran porte de los dinosaurios y el comparativamente pequeño tamaño de los mamíferos modernos ha sido cuantificado por Nicholas Hotton. Basándose en 63 géneros de dinosaurios, los datos de Hotton muestran una masa promedio que excede los 395.9 kg (el peso de un oso grizzly mediano), y una masa genérica mediana de cerca de dos toneladas (comparable a una jirafa). Esto contrasta marcadamente con mamíferos extintos (788 géneros) cuya masa promedio es de 631 gramos (la de un roedor pequeño). El dinosaurio no aviano más pequeño hallado hasta hoy era mayor que dos tercios de todos los mamíferos actuales; la mayoría de los dinosaurios eran mayores que todos los mamíferos vivientes con excepción de un 2 % de los individuos.[135]
Los saurópodos fueron los dinosaurios más grandes y pesados. Durante la mayor parte de la era de los dinosaurios, incluso los menores saurópodos eran más grandes que cualquier otro animal en sus hábitats, y los mayores eran por un orden de magnitud más masivos que cualquier otra criatura que haya caminado sobre la Tierra. Los mamíferos prehistóricos gigantes tales como Paraceratherium (el mayor mamífero terrestre conocido) eran enanos junto a los saurópodos gigantes, y solo las ballenas modernas alcanzan o sobrepasan a los saurópodos en tamaño.[136] Hay varias ventajas propuestas para el gran tamaño de los saurópodos, incluyendo la protección de la depredación, la reducción en el uso de energía y la longevidad, pero puede que la más importante de ellas tuviera relación con su dieta. Los animales grandes son más eficientes digiriendo que los pequeños, ya que la comida pasa más tiempo en sus sistemas digestivos. Esto también les permite subsistir con comida de valor nutritivo más bajo que los animales pequeños. Los restos de saurópodos son hallados principalmente en formaciones rocosas que son interpretadas como de ambientes secos o secos estacionalmente, por lo que la habilidad de comer grandes cantidades de plantas con pocos nutrientes hubiera sido ventajosa en esa clase de ecosistemas.[137]
Grandes y pequeños
[editar]Probablemente los científicos nunca sabrán cuales fueron los dinosaurios más grandes y más pequeños que hayan existido. Esto se debe a que solo un pequeño porcentaje de los animales llega a fosilizarse, y muchos de estos permanecen enterrados. Pocos de los especímenes que se recuperan son esqueletos completos y las impresiones de piel y otros tejidos blandos son muy raras. Reconstruir un esqueleto completo a través de comparar el tamaño y la morfología de los huesos de las especies mejor conocidas y que más se asemejen es un arte inexacto, y la reconstrucción de los músculos y otros órganos de la criatura viva es, como mucho, un proceso de establecer suposiciones fundamentadas.[138]

El dinosaurio más alto y pesado conocido de esqueletos completos es Giraffatitan brancai (previamente clasificado como una especie de Brachiosaurus). Sus restos fueron descubiertos en Tanzania entre 1907 a 1912. Huesos de varios individuos de tamaño parecido fueron incorporados en el esqueleto ahora montado y expuesto en el Museo de Historia Natural de Berlín;[139] este esqueleto mide 12 m de alto y entre 21.8 y 22.5 m de largo,[140][141] y le pertenecería a un animal que pesaría entre 30 000 y 60 000 kg (kilogramos). El dinosaurio completo más largo es un espécimen de 27 m de largo de un Diplodocus, el cual fue descubierto en Wyoming en los Estados Unidos y se expone en el Museo Carnegie de Historia Natural en Pittsburgh desde 1907.[142]
Hubo dinosaurios mucho mayores, pero el conocimiento de estos se basa en un pequeño número de fósiles fragmentarios. Muchos de los mayores especímenes registrados de herbívoros fueron todos descubiertos en la década de 1970 en adelante, e incluyen al enorme Argentinosaurus, el cual pudo haber pesado entre 80 000 y 100 000 kg; algunos de los más largos incluyen a Diplodocus hallorum de 33,5 m[137] (antes conocido como Seismosaurus) y a Supersaurus de 33 m de largo;[143] y entre los más altos, se encuentra a Sauroposeidon con 18 m de altura, el cual pudo haber alcanzado la ventana de un sexto piso. El más pesado y largo de todos ellos pudo haber sido Amphicoelias fragillimus, conocido solo por una vértebra parcial (el arco neural) descrita en 1878 y a la fecha diciembre de 2016 desaparecida. Extrapolando a partir de la ilustración de este hueso, el animal pudo haber medido 58 m de largo y pesado unos 120 000 kg.[137] El mayor dinosaurio carnívoro conocido fue Spinosaurus, alcanzando una longitud de más de 16 metros, y con un peso de 8150 kg.[144]
Otros grandes dinosaurios carnívoros incluyen a los terópodos Giganotosaurus, Carcharodontosaurus y Tyrannosaurus.[145] Therizinosaurus y Deinocheirus estuvieron entre los más altos entre los terópodos.
Sin incluir a las aves (Avialae), los dinosaurios más pequeños conocidos eran del tamaño de palomas.[146] No resulta sorprendente que los dinosaurios no aviares más pequeños fueran aquellos más cercanamente relacionados con las aves. Anchiornis huxleyi, por ejemplo, tenía un esqueleto cuya longitud total era de menos de 35 cm.[146][147] A. huxleyi es reconocido como el dinosaurio no aviano más pequeño descrito de un espécimen adulto, con un peso estimado de 110 g.[147] Los dinosaurios herbívoros no aviano más pequeños incluyen a Microceratus y a Wannanosaurus, con cerca de 60 cm de longitud cada uno.[12][148]
Comportamiento
[editar]
Muchas aves modernas son muy sociales, viviendo con frecuencia en bandadas. Hay un consenso generalizado de que algunos comportamientos que son comunes en las aves, así como en los cocodrilos (los parientes vivos más cercanos de las aves), eran también comunes entre los grupos extintos de dinosaurios. Las interpretaciones del comportamiento de las especies animales conocidas por restos fósiles se basan generalmente en la postura de sus esqueletos y su hábitat, las simulaciones por computadora de su biomecánica, y las comparaciones con animales modernos que tengan nichos ecológicos similares.[128]
La primera evidencia de que el comportamiento en manada era común en varios grupos de dinosaurios aparte de las aves se produjo en 1878 con el descubrimiento de 31 ejemplares de Iguanodon bernissartensis, ornitisquios que se piensa perecieron juntos en un yacimiento en Bernissart, Bélgica, tras haber caído en un profundo socavón inundado en que se ahogaron.[149] Otros sitios de muerte en masa fueron descubiertos posteriormente. Estos, junto con varias huellas fosilizadas, sugieren que el comportamiento gregario era común en varias especies de dinosaurios. Las huellas de cientos o incluso miles de herbívoros indica que los hadrosáuridos pueden haberse movido en grandes grupos, como ocurre con el bisonte americano o la gacela saltarina. Las huellas de saurópodos documentan que estos animales viajaban en grupos compuestos de diferentes especies, al menos en Oxfordshire, Inglaterra,[150] aunque esto no es evidencia de una estructura específica de las manada.[151] El congregarse en grupos pudo haber evolucionado para defenderse, para propósitos de migración, o para proporcionar protección a las crías. Hay evidencia de que muchos tipos de dinosaurios de crecimiento lento, incluyendo a varios terópodos, saurópodos, anquilosaurios, ornitópodos y ceratopsios, formaban agregaciones de individuos inmaduros. Un ejemplo es un sitio en Mongolia Interior en que se han hallado los restos de cerca de 20 Sinornithomimus, de entre uno a siete años de edad. Se ha interpretado que esta asociación corresponde a un grupo social que quedó atrapado en el fango.[152] La interpretación de los dinosaurios como animales sociales también se ha extendido a los terópodos carnívoros, representándolos como cazadores grupales que cooperaban para derribar presas de gran tamaño.[153][154] Sin embargo, este estilo de vida no es común entre las aves modernas, cocodrilos y otros reptiles, y la evidencia tafonómica sugiere que la supuesta cacería en grupo al estilo de los mamíferos sugerida para dinosaurios como Deinonychus y Allosaurus puede interpretarse también como el resultado de disputa fatales entre animales que se encontraban alimentándose, como se ha observado en varios diápsidos depredadores modernos.[155]

Las crestas y golas de algunos dinosaurios, como los marginocéfalos, terópodos y lambeosaurinos, pueden haber sido demasiado frágiles para haber sido usadas como defensa activa, y por tanto probablemente eran usadas para exhibiciones sexuales o de agresión, aunque poco se sabe sobre el apareamiento y el territorialismo en los dinosaurios. Heridas en la cabeza por mordidas sugieren que por lo menos los terópodos luchaban en confrontaciones agresivas.[156]
Desde el punto de vista del comportamiento, uno de los más valiosos fósiles de dinosaurios fue el hallado en el desierto de Gobi en 1971. Este incluye a un Velociraptor atacando a Protoceratops,[157] lo cual proporciona evidencia de que los dinosaurios ciertamente se atacaban entre sí.[158] Evidencia adicional de ataques a presas vivas se halla en la cola parcialmente curada de un Edmontosaurus, un dinosaurio hadrosáurido; la cola fue dañada de tal manera que muestra que el animal fue atacado por un tiranosáurido pero logró sobrevivir.[158] El canibalismo entre algunas especies de dinosaurios fue confirmado por marcas de dientes halladas en Madagascar en 2003, en el terópodo Majungasaurus.[159]
Las comparaciones realizadas con los anillos escleróticos de dinosaurios, aves y reptiles modernos han sido utilizadas para inferir los patrones de actividad diaria de los dinosaurios. Aunque se ha sugerido que la mayoría de los dinosaurios eran activos durante el día, estas comparaciones han mostrado los pequeños dinosaurios depredadores como los dromeosáuridos, Juravenator y Megapnosaurus eran probablemente nocturnos. Los dinosaurios herbívoros y omnívoros de tamaño medio y grande como los ceratopsios, sauropodomorfos, hadrosáuridos y ornitomimosaurios pueden haber sido catemerales, es decir activos durante intervalos cortos a lo largo del día, aunque se infiere que el pequeño ornitisquio Agilisaurus hubiera sido diurno.[160]
Basándose en la evidencia fósil disponible de dinosaurios como Oryctodromeus, parece que algunas especies de ornistisquios parecen haber adoptado un estilo de vida parcialmente fosorial (en madrigueras).[161] Muchas aves modernas son arborícolas, y esto era igual para muchas aves del Mesozoico, especialmente las enantiornites.[162] Aunque algunas especies de dinosaurios similares a aves parecen también haber sido arborícolas (incluyendo a dromeosáuridos tales como Microraptor [163]) muchos dinosaurios no avianos parecen haber sido ante todo de locomoción terrestre. El buen entendimiento de como los dinosaurios se desplazaban en tierra es clave para modelar el comportamiento dinosauriano; la ciencia de la biomecánica, en particular, ha provisto un significativo entendimiento en esta área. Por ejemplo, los estudios de las fuerzas ejercidos por los músculos y la gravedad en la estructura esquelética de los dinosaurios han indagado cuan rápido podían correr los dinosaurios,[164] si los diplodócidos podrían crear boom sónicos a través de agitar sus colas parecidas a látigos,[165] y si los saurópodos podían flotar.[166]
Comunicación
[editar]
Las aves actuales son bien conocidas por comunicarse usando principalmente señales visuales y auditivas, y la amplia diversidad de estructuras de exhibición visual entre los grupos extintos de dinosaurios sugiere que la comunicación visual fue siempre importante en la biología dinosauriana. Por otro lado, se sabe menos sobre la vocalización en los dinosaurios. En 2008, el paleontólogo Phil Senter examinó la evidencia de vocalización en la vida animal del Mesozoico, incluyendo a los dinosaurios.[167] Senter encontró que, a diferencia de las representaciones populares de dinosaurios rugiendo de las películas, es posible que muchos dinosaurios mesozoicos no fueran capaces de crear ningún tipo de vocalizaciones (si bien las crestas huecas de los lambeosaurinos pudieron funcionar como cámaras de resonancia usadas para un amplio rango de sonidos).[168][169] Para llegar a esta conclusión, Senter estudió la distribución de los órganos vocales en aves y reptiles modernos. Él halló que las cuerdas vocales en la laringe probablemente evolucionaron varias veces entre los reptiles, incluyendo a los crocodilianos, los cuales son capaces de producir rugidos guturales. Las aves, por otro lado, carecen de laringe. En cambio, sus sonidos los hacen con la siringe, un órgano vocal solo hallado en las aves, y que no está relacionado con la laringe, lo que significa que evolucionó independientemente de los órganos vocales de los reptiles. La siringe depende del sistema aviano de sacos aéreos para funcionar; específicamente, requiere de la presencia de un saco aéreo clavicular cerca de la fúrcula. Este saco aéreo deja marcas o aberturas distintivas en los huesos, incluyendo un agujero característico en el hueso superior del brazo (húmero). Aunque el extenso sistema de sacos aéreos es una característica única de los dinosaurios saurisquios, el saco aéreo clavicular que es necesario para vocalizar no aparece en el registro fósil hasta las aves enantiornites (con la excepción de Aerosteon, que probablemente evolucionó su saco aéreo clavicular independientemente de las aves por razones distintas a la vocalización).[167]
Los dinosaurios más primitivos con una siringe para hacer sonidos son las aves enantiornites, y por tanto los arcosaurios de la línea aviana (Ornithodira) no realizaban llamadas vocales. En cambio, varias líneas de evidencia sugieren que los dinosaurios usaban principalmente la comunicación visual, a través de cuernos, golas, crestas, velas y plumas de apariencia distintiva (y posiblemente de colores brillantes). Esto es similar a lo que se observa en algunos reptiles modernos tales como los lagartos, que en su mayor parte son silenciosos (aunque como muchos dinosaurios, estos poseen un sentido del oído bien desarrollado) pero usan comportamientos de exhibición complejos y colores llamativos para comunicarse.[167]
Además, los dinosaurios pudieron usar otros métodos para producir sonidos a fin de comunicarse. Otros animales, reptiles incluidos, usan una amplia variedad de sonidos no vocales, incluyendo el siseo, golpes o chirridos con la mandíbula, el uso de elementos en el ambiente (como el chapoteo), y batir las alas (algo posible en los dinosaurios manirraptores alados).[167]
Biología reproductiva
[editar]
Todos los dinosaurios depositan huevos amnióticos con cáscaras duras compuestas principalmente de carbonato de calcio.[170] Los huevos son usualmente dejados en un nido. Muchas especies crean nidos elaborados, en forma de cuenco, domo, meseta, camas, montículos, o madrigueras.[171] Algunas especies de aves modernas no hacen nidos; una especie que vive en acantilados, el arao común deja sus huevos en la roca desnuda, y el macho del pingüino emperador cuida de sus huevos colocándolos entre su vientre y sus patas. Las aves primitivas y muchos de los dinosaurios no aviares con frecuencia depositaban sus huevos en nidos comunales, en los que los mayormente los machos incubaban los huevos. Aunque las aves modernas solo tengan un oviducto funcional y depositen un huevo a la vez, las aves más primitivas y los dinosaurios tenían dos, como los cocodrilos. Algunos dinosaurios no aviares, como Troodon, presentaban puestas repetitivas, en las que los adultos podían depositar un par de huevos cada día o dos, y luego se aseguraban de que eclosionaran simultáneamente al posponer su incubación hasta que todos los huevos hubieran sido depositados.[172]
Cuando depositaban los huevos, las hembras desarrollaban un tipo especial de hueso entre la capa ósea dura externa y la médula de sus extremidades. Este hueso medular, el cual es rico en calcio, es usado para hacer la cáscara del huevo. El descubrimiento de esta característica en un espécimen de Tyrannosaurus rex proporcionó evidencia del hueso medular en los dinosaurios extintos y, por primera vez, permitió a los paleontólogos establecer el sexo de un ejemplar fósil de dinosaurio. Investigaciones posteriores han encontrado hueso medular en el carnosaurio Allosaurus y el ornitópodo Tenontosaurus. Debido a que el linaje de dinosaurios que incluye a Allosaurus y a Tyrannosaurus se separó del que conduce a Tenontosaurus muy tempranamente en la evolución de los dinosaurios, esto sugiere que la producción de tejido medular es una característica general de todos los dinosaurios.[173]

Otro rasgo extendido entre las aves modernas es el cuidado paternal para las crías recién nacidas. Jack Horner en 1978 descubrió una zona de anidación de Maiasaura (el "lagarto buena madre") en Montana, Estados Unidos, demostrando que el cuidado paternal continuaba por un buen tiempo después del nacimiento entre los ornitópodos, lo que indica que este comportamiento puede haber sido común entre los dinosaurios.[174] Hay evidencia de que otros dinosaurios no terópodos, como los titanosaurios de la Patagonia también anidaban en grandes grupos.[175] Un espécimen hallado en Mongolia del ovirraptórido Citipati osmolskae fue descubierto en una postura de incubación similar a la de las gallinas en 1993,[176] lo cual indica que usaban una capa aislante de plumas para mantener el calor de los huevos.[177] La suposición de que el cuidado paternal fuera un rasgo común en los dinosaurios es apoyado por otros hallazgos. Por ejemplo, un embrión de dinosaurio perteneciente al prosaurópodo Massospondylus fue encontrado sin dientes, indicando que se requería que las crías fueran alimentadas por sus padres.[178] Los senderos de huellas también han confirmado cuidado paternal entre ornitópodos de la Isla de Skye en el noroeste de Escocia.[179] Nidos y huevos han sido hallados para varios de los principales grupos de dinosaurios, y parece probable que todos los dinosaurios cuidaran de sus crías en alguna medida antes o poco después de la eclosión.[180] Recientemente se han reconocido, gracias a los estudios del ámbar, la relación estrecha de algunos grupos de dinosaurios emplumados con artrópodos que vivirían en el interior de sus nidos. Así se conocen garrapatas[181] que se alimentarían de la sangre de los dinosaurios, y escarabajos derméstidos[182] que comerían sus restos de plumas y piel.
Fisiología
[editar]Debido a que tanto los crocodilianos como las aves modernas tienen corazones divididos en cuatro cámaras (si bien modificados en los crocodilianos), es probable que sean un rasgo compartido por todos los arcosaurios, incluyendo a los dinosaurios.[183] Aunque todas las aves tienen metabolismos altos y son de "sangre caliente" (endotérmico), ha existido un vigoroso debate desde la década de 1960 con respecto a que tanto se remonta esta característica en el linaje dinosauriano. Los científicos han diferido en si los dinosaurios no aviares eran endotérmicos, ectotérmicos o una combinación de ambas cosas.[184]
Tras el descubrimiento de los primeros dinosaurios no aviares, los paleontólogos inicialmente consideraron que eran ectotérmicos. Esta supuesta condición de "sangre fría" fue usada para implicar que los dinosaurios eran organismos relativamente lentos y perezosos, aunque muchos reptiles modernos en realidad son rápidos y ágiles a pesar de depender de fuentes externas de calor para regular su temperatura corporal. La idea de los dinosaurios lentos continuó siendo la predominante hasta que Robert T. «Bob» Bakker, uno de los primeros en proponer la endotermia de los dinosaurios, publicó un influyente artículo sobre el tema en 1968.[185]
La evidencia reciente indica que incluso los dinosaurios no aviares y las aves prosperaron en climas fríos, y al menos algunas de las primeras especies deben de haber sido capaces de regular su temperatura corporal por medio biológicos internos (ayudado por la masa de los animales en las especies más grandes y las plumas u otros elementos aislantes en las especies más pequeñas). La evidencia de endotermia en los dinosaurios mesozoicos incluyen el descubrimiento de los «dinosaurios polares» en Australia y Antártida así como el análisis de estructuras de vasos sanguíneos dentro de los huesos fósiles que son típicos de los endotermos. El debate científico ha continuado en lo que respecta a los modos específicos en que evolucionó la regulación de temperatura en los dinosaurios.[186]

En los dinosaurios saurisquios, los metabolismos activos son apoyados por la evolución del sistema respiratorio aviano, caracterizado por un extenso sistema de sacos aéreos que se extiende a los pulmones e invade a muchos de los huesos en el esqueleto, lo que lo hace más ligero.[187] Los primeros sistemas respiratorios de tipo aviano con sacos aéreos pueden haber sido capaces de sostener mayores niveles de actividad que en los mamíferos de tamaño y constitución similar. Además de proveer un muy eficiente suministro de oxígeno, el rápido flujo de aire pudo haber sido un efectivo mecanismo de enfriamiento, lo cual es esencial en animales que son activos pero demasiado grandes para disipar todo el exceso de calor a través de su piel.[188]
Como otros reptiles, los dinosaurios son principalmente uricotélicos, lo que significa que sus riñones extraen los desechos nitrogenados de su flujo sanguíneo y lo excretan como ácido úrico en lugar de la urea o amoníaco como en otros animales, a través de los uréteres a los intestinos. En muchas especies actuales, el ácido úrico es excretado junto con las heces como un desecho semisólido.[189][190][191] Sin embargo, al menos algunas aves (como los colibríes) pueden ser facultativamente amoniotélicas, excretando la mayor parte de los desechos nitrogenados como amoníaco.[192] Estas también excretan creatina, en lugar de creatinina como en los mamíferos. Este material, así como el desecho de los intestinos, sale por la cloaca.[193][194] Aparte de esto, muchas especies regurgitan egagrópilas, y se conocen fósiles de egagrópilas posiblemente procedentes de dinosaurios que se remontan hasta el período Cretácico.[195]
Enfermedades
[editar]- En agosto de 2020, se descubrió por primera vez un cáncer maligno en un dinosaurio. El hueso canceroso en el que se descubrió fue el peroné de un Centrosaurus apertus, un dinosaurio con cuernos que vivió hace entre 76 y 77 millones de años. [2]
Origen de las aves
[editar]La posibilidad de que los dinosaurios fueran los antepasados de las aves fue sugerida por primera vez en 1868 por Thomas Henry Huxley.[196] Después del trabajo de Gerhard Heilmann a principios del siglo XX, la teoría de las aves como descendientes de los dinosaurios fue abandonada en favor de antepasados tecodontes, siendo la prueba clave la supuesta falta de clavícula de los dinosaurios.[197] Sin embargo, como lo demostraron descubrimientos posteriores, las clavículas (o una única fúrcula fusionada, derivada de dos clavículas separadas) no estaban realmente ausentes;[99] ya se había encontrado en 1924 en Oviraptor, pero habían sido identificadas erróneamente como interclavícula.[198] En la década de 1970, John Ostrom recuperó la teoría de los dinosaurios como antepasados de las aves,[199] que ganó impulso durante las décadas siguientes con el advenimiento del análisis cladístico,[200] y un gran aumento de los descubrimientos de pequeños terópodos y aves primitivas.[37] Han sido de especial relevancia los fósiles de la formación de Yixian, donde se han encontrado múltiples terópodos y aves primitivas, muchas veces con plumas de algún tipo.[99] Las aves comparten más de un centenar de rasgos anatómicos distintos con los dinosaurios terópodos, al siglo XXI considerados generalmente sus parientes prehistóricos más cercanos.[201]
El grupo más cercano a ellos es el de los celurosaurios manirraptores.[99] El origen dinosauriano de las aves no solo esta respaldado por la evidencia paleontológica sino también por las secuencias proteicas obtenidas de las proteínas de Tyrannosaurus rex y Brachylophosaurus canadensis que demuestran que están emparentados con las aves, aunque estos dos últimos pueden estar más emparentados entre ellos que con las aves. La filogenia dio el siguiente resultado:[202][203]
| Archosauria |
| ||||||||||||||||||

Una minoría de científicos, notablemente Alan Feduccia y Larry Martin, han sugerido otros caminos evolutivos, incluyendo versiones revisadas de la propuesta de Heilmann de los arcosaurios basales,[204] o que los terópodos manirraptores eran los antepasados de las aves, pero que no eran dinosaurios, sino convergentes con ellos.[11]
Plumas
[editar]
Las plumas son una de las características más reconocibles de las aves modernas, y un rasgo que fue compartida por todos los otros grupos de dinosaurios. Basado en la distribución actual de la evidencia fósil, al parecer las plumas fueron un rasgo dinosauriano ancestral, aunque uno que se puede haber perdido de forma selectiva en algunas especies.[205] Han sido descubiertas evidencias fósil directa de plumas o de estructuras similares a plumas en una gran variedad de especies en muchos grupos de dinosaurios no aviares, tanto entre los saurisquios como los ornitisquios. Las estructuras simples y ramificados, como plumas se conocen de heterodontosáuridos, neornitisquianos primitivos[206] terópodos,[207] y ceratópsidos primitivos. Solo se ha encontrado evidencias de plumas verdaderas, paletas similares a las plumas de vuelo de las aves modernas en el subgrupo terópodo Maniraptora que incluye oviraptorosaurios, troodóntidos dromeosáuridos, y aves.[99][208] Las estructuras similares a plumas conocidas como pycnofibras también se han encontrado en los pterosaurios,[209] lo que sugiere la posibilidad de que filamentos similares a plumas podrían haber sido comunes en el linaje de aves y evolucionado antes de la aparición de los propios dinosaurios.[205] La investigación sobre la genética de los cocodrilos americanos también ha revelado que los escudos cocodrilianos sí poseen plumas-queratinas durante el desarrollo embrionario, pero estas queratinas no son expresadas por los animales antes de la eclosión.[210]
Archaeopteryx fue el primer fósil encontrado que reveló una posible relación entre los dinosaurios y las aves. Se considera un fósil de transición, en que se muestra características de ambos grupos. Sacado a la luz dos años después seminal de Darwin El origen de las especies, su descubrimiento estimuló el debate incipiente entre los partidarios de la biología evolutiva y el creacionismo. Esta ave es tan antigua como lo son los dinosaurios, sin una clara impresión de plumas en la roca circundante, al menos un espécimen fue confundido con Compsognathus.[211] Desde los años 1990 se ha encontrado un número creciente de dinosaurios no aviares con plumas, lo cual proveyó nuevas pruebas adicionales sobre la relación directa existente entre dinosaurios y aves modernas. La mayoría de estos especímenes han sido exhumados en la provincia de Liaoning en el noreste de China, que fue parte de un continente aislado durante el Cretáceo. A pesar de que rastros de plumas han sido encontrados solo en la formación Yixian y otros pocos sitios, es probable que dinosaurios similares en otras partes del mundo también hayan estado cubiertos por plumas. La falta de restos fósiles de plumaje a escala planetaria puede deberse al hecho de que las estructuras delicadas como piel, pelo y plumas no son preservadas por las usuales condiciones de fosilización, lo que dificulta su presencia en el registro fósil.[212]
La descripción de los dinosaurios emplumados no ha estado exento de polémica; tal vez los mayores críticos han sido Alan Feduccia y Theagarten Lingham-Soliar, que han propuesto que algunos fósiles similares a plumas supuestamente son el resultado de la descomposición de las fibras de colágeno que está debajo de la piel de los dinosaurios,[213][214][215] y que los dinosaurios maniraptoranos con plumas paletas no eran en realidad dinosaurios, sino convergentes con los dinosaurios.[11][214] Sin embargo, sus opiniones en general no han sido aceptadas por otros investigadores, hasta el punto de ser cuestionada la naturaleza científica de las propuestas de Feduccia.[216]
Esqueleto
[editar]Como las plumas están generalmente asociadas a las aves, los dinosaurios emplumados son tildados y descritos con frecuencia como el eslabón perdido entre las aves y los dinosaurios. Sin embargo, las múltiples características esqueléticas que comparten los dos grupos también son un indicio importante para los paleontólogos. Las áreas del esqueleto con semejanzas importantes son el cuello, el pubis, la muñeca (carpo semilunar), el brazo y la cintura escapular, la fúrcula y la quilla. La comparación de los esqueletos de dinosaurio y de las aves por medio del análisis cladístico refuerza la teoría de la relación entre ambos grupos.[217]
Tejidos blandos
[editar]
Los grandes dinosaurios carnívoros poseían un complejo sistema de sacos de aire similares a los que tienen las aves modernas, según una investigación de 2005 dirigida por Patrick O'Connor de la Universidad de Ohio. Es probable que los pulmones de los dinosaurios terópodos carnívoros bípedos con patas (similares a las de las aves) bombeen aire en sacos vacíos de su esqueleto, como ocurre en las aves. Lo que en el pasado se consideraba formalmente un rasgo único de las aves estaba presente en cierta forma en los antepasados de estas, dijo O'Connor.[218] En 2008, los científicos describieron a Aerosteon riocoloradensis, el esqueleto del que ha proporcionado las pruebas más claras hasta ahora de un dinosaurio con un sistema de respiración similar a la de las aves. Una tomografía computarizada puso de manifiesto la presencia de sacos de aire dentro de la cavidad corporal del esqueleto de Aerosteon.[219][220]
Evidencias de comportamiento
[editar]Los fósiles de troodóntidos Mei y Sinornithoides demuestran que algunos dinosaurios dormían con la cabeza bajo el brazo.[221] Este comportamiento, que pueden haber contribuido a mantener la cabeza caliente, también es característico de las aves modernas. Varios especímenes deynonsaurianos y oviraptorosaurios también se han encontrado conservados en la parte superior de sus nidos, rumiando probablemente de una manera similar a las aves.[222] La relación entre el volumen del huevo y la masa corporal de los adultos entre estos dinosaurios sugieren que los huevos son incubados principalmente por el macho, y que los jóvenes fueron muy precoces, similares a muchas aves modernas que habitan en el suelo.[223]
Algunos dinosaurios se sabe que usan piedras de molleja, como las aves modernas. Estas piedras son ingeridas por animales para ayudar a la digestión y descomponer los alimentos y fibras duras una vez que entran en el estómago. Cuando se encuentra en asociación con fósiles, estas piedras se llaman gastrolitos.[224]
Extinción de grupos mayores
[editar]El descubrimiento de que las aves son un tipo de dinosaurio demostró que en general los dinosaurios no están, de hecho, extintos como se dice habitualmente.[225] Sin embargo, todos los dinosaurios no aviares, así como muchos grupos de aves se extinguieron repentinamente hace aproximadamente 66 millones de años. Se ha sugerido que debido a que los pequeños mamíferos, squamatas y aves ocuparon los nichos ecológicos aptos para su tamaño corporal pequeño, los dinosaurios no aviares nunca evolucionaron una diversa fauna de especies de cuerpo pequeño, lo que condujo a su caída, cuando se vieron afectados por los tetrápodos terrestres de gran cuerpo, como consecuencia del evento de extinción masiva.[226] Muchos otros grupos de animales también se extinguieron en este momento, incluyendo las amonitas (moluscos similares a nautilus), mosasaurios, plesiosaurios, pterosaurios, y muchos grupos de mamíferos.[15] De manera significativa, los insectos no sufrieron ninguna pérdida apreciable en su población, lo que dejó alimento a disposición para otros sobrevivientes. Esta extinción masiva se la conoce como extinción masiva del Cretácico-Paleógeno. La naturaleza del evento que causó esta extinción en masa ha sido ampliamente estudiada desde la década de 1970; al siglo XXI, varias teorías relacionadas cuentan con el apoyo de los paleontólogos. Aunque el consenso es que un evento de impacto fue la causa principal de la extinción de los dinosaurios, algunos científicos citan otras causas posibles, o apoyan la idea de que una confluencia de varios factores fue responsable de la repentina desaparición de los dinosaurios del registro fósil.[227][228][229]
En el punto álgido del Mesozoico, no había casquetes polares, y el nivel del mar era entre 100 y 250 m más alto que en la actualidad. La temperatura del planeta también era mucho más uniforme, con una diferencia de solo 25 °C (45 °F) entre las temperaturas polares y las ecuatoriales. En promedio, las temperaturas atmosféricas también eran mucho más cálidas; los polos, por ejemplo, eran 50 °C (90 °F) más cálidos que hoy en día.[230][231]
La composición atmosférica durante el Mesozoico también era muy diferente. Los niveles de dióxido de carbono eran hasta doce veces superiores a los de la actualidad, y el oxígeno representaba entre el 32-25 % de la atmósfera, en comparación con el 21 % de hoy en día. Sin embargo, a finales del Cretácico, el ambiente estaba cambiando dramáticamente. Se redujo la actividad volcánica, provocando una tendencia de enfriamiento a medida que los niveles de dióxido de carbono atmosférico caían. Los niveles de oxígeno en la atmósfera también comenzaron a fluctuar y acabaron cayendo considerablemente. Algunos científicos creen que el cambio climático, junto con los niveles más bajos de oxígeno, podría haber provocado directamente la desaparición de muchas especies. Si los dinosaurios tenían sistemas respiratorios similares a los típicos de las aves modernas, podría haber sido particularmente difícil para ellos adaptarse a un rendimiento respiratorio inferior, teniendo en cuenta la enorme cantidad de oxígeno que necesitaban sus cuerpos.[15]
Colisión de un asteroide
[editar]
La teoría de la colisión con un asteroide, que fue popularizada en 1980 por Walter Alvarez y sus colegas, relaciona la extinción de finales del período Cretácico con un impacto con un bólido hace aproximadamente 65,5 millones de años. Alvarez y otros sugirieron que un aumento repentino de los niveles de iridio, registrado en todo el mundo en los estratos rocosos de este periodo, era una prueba directa del impacto.[232] La mayoría de los indicios sugieren a 2016 que un bólido de entre 5 y 15 km de diámetro impactó con la Tierra cerca de la península de Yucatán, creando el cráter de Chicxulub, de 170 km de diámetro, y desencadenando la extinción masiva.[233] Los científicos no saben con certeza si los dinosaurios estaban prosperando o en declive antes del impacto astronómico. Algunos científicos sugieren que el meteorito causó una bajada de la temperatura atmosférica larga y no natural, mientras que otros, en cambio, afirman que habría provocado una ola de calor fuera de lo habitual. Algunos científicos proponen que el impacto del meteorito provocó extinciones tanto directamente (por el calor del impacto meteórico) como indirectamente (por un enfriamiento global debido al hecho de que la materia eyectada del cráter de impacto reflejaba la radiación térmica del Sol). Aunque la velocidad de extinción no puede ser inferida solo del registro fósil, varios modelos sugieren que la extinción fue extremadamente rápida, siendo de horas en lugar de años.[234]
Traps del Decán
[editar]Antes de 2000, los argumentos que las Traps del Decán causaron la extinción estaban generalmente relacionados con la teoría de que la extinción fue gradual, pues se pensaba que las Traps comenzaron hace unos sesenta y ocho millones de años y duraron unos dos millones. Sin embargo, hay pruebas de que dos tercios de las Traps del Decán fueron creadas en un período de un millón de años, hace aproximadamente 65,5 millones de años, por lo que estas erupciones habrían provocado una extinción bastante rápida, posiblemente en un período de miles de años, pero igualmente un período más largo que el que cabría esperar de un único impacto meteórico.[235][236]
Las Traps del Decán podrían haber causado la extinción mediante diversos mecanismos, incluyendo la liberación de polvo y de aerosoles sulfúricos en el aire, que podrían haber bloqueado la luz solar, reduciendo la capacidad fotosintética de las plantas. Además, el vulcanismo de las Traps del Decán podría haber provocado emisiones de dióxido de carbono que habrían aumentado el efecto invernadero una vez el polvo y los aerosoles hubieran desaparecido de la atmósfera.[236] Antes de la extinción masiva de los dinosaurios, la liberación de gases volcánicos durante la formación de las Traps del Decán contribuyeron a un calentamiento global aparentemente masivo. Algunos datos indican un aumento medio de la temperatura de aproximadamente 8 °C (grados Celsius) en el último medio millón de años antes del impacto en Chicxulub.[235][236]
En los años en que la teoría de las Traps del Decán estaba relacionada con una extinción más lenta, Luis Walter Álvarez (que murió en 1988), respondía que los paleontólogos estaban siendo engañados por los pocos datos de que disponían. Aunque en principio su afirmación no fue bien recibida, estudios de campo intensivos posteriores de los estratos fosilíferos reforzaron su argumento. Finalmente, la mayoría de paleontólogos comenzaron a aceptar la idea de que las extinciones en masa de finales del Cretáceo se debieron principalmente, o al menos en parte, a un impacto masivo de la Tierra contra un bólido. Sin embargo, incluso Walter Alvarez admitió que hubo otros grandes cambios en la Tierra incluso antes del impacto, tales como un descenso del nivel del mar y grandes erupciones volcánicas que crearon las Traps del Decán en la India, y que estos cambios podrían haber contribuido a las extinciones.[237]
Falta de adaptación a las condiciones cambiantes
[editar]El Cretácico medio, las plantas con flor se convirtieron en una parte importante de los ecosistemas terrestres, que anteriormente habían sido dominados por gimnospermas como las coníferas.[238] Los coprolitos (estiércol fosilizado) de dinosaurio indican que, mientras que algunos comían angiospermas, la mayoría de dinosaurios herbívoros se alimentaban principalmente de gimnospermas. Análisis estadísticos de Lloyd et al. llegaron a la conclusión de que, a diferencia de lo que afirmaban estudios anteriores, los dinosaurios no se diversificaron mucho en el Cretáceo superior. Lloyd et al. sugirieron que el fracaso de los dinosaurios a la hora de adaptarse a ecosistemas cambiantes los condenó a la extinción.[239]
Posibles supervivientes del Paleoceno
[editar]Ocasionalmente se encuentran restos de dinosaurios no aviares en sedimentos por encima del límite K-T. En 2000, Fassett y colaboradores comunicaron el descubrimiento del fósil de un hueso de una pata de un hadrosaurio en la cuenca del San Juan (Nuevo México), y el describieron este único hallazgo como prueba de dinosaurios del Paleoceno. La formación en la que se descubrió el hueso fue datada de principios de la época del Paleoceno, hace aproximadamente 64,5 millones de años. Si el hueso no fue redepositado en ese estrato por la acción de la reelaboración de materiales más antiguos, sería una prueba de que algunas poblaciones de dinosaurios podrían haber sobrevivido al menos medio millón de años en la era Cenozoico.[240] Otras pruebas incluyen el descubrimiento de restos de dinosaurios en la formación de Hell Creek hasta 1,3 m por encima del límite K-T (equivalentes a 40 000 años después).[241] Han llegado descubrimientos similares de otros lugares del mundo, incluyendo China. Sin embargo, muchos científicos rechazan que hubiera habido dinosaurios en el Paleoceno, argumentando que estos huesos fueron reelaborados, es decir, que la erosión los sacó de su ubicación inicial y quedaron enterrados de nuevo en sedimentos posteriores,[242][243] o piensan que, aunque fuera cierto, la presencia de un grupo de dinosaurios en el Paleoceno inferior no cambiaría gran cosa a efectos prácticos.[242]
En la cultura popular
[editar]

Según la visión generalizada, los dinosaurios eran criaturas de apariencia fantástica y a menudo de un enorme tamaño. Así pues, han atraído la imaginación del público y se han convertido en una parte duradera de la cultura. Solo tres décadas después de las primeras descripciones científicas de restos de dinosaurios, se les erigieron unas esculturas en el Crystal Palace Park de Londres que gustaron tanto al público que se vendieron réplicas a pequeña escala lo que se convirtió en uno de los primeros ejemplos de merchandising de productos derivados. En las décadas posteriores, se han hecho exhibiciones de dinosaurios en parques y museos de todo el mundo, respondiendo al interés público y reforzándolo.[244] Desde hace tiempo la popularidad de los dinosaurios también ha tenido un efecto recíproco sobre el conocimiento de los dinosaurios. La competición entre los museos para la atención del público llevó directamente a la Guerra de los Huesos de las décadas de 1880 y 1890, y la contribución resultante al conocimiento de estos animales fue enorme.[245]
Los dinosaurios ocupan un lugar importante en la cultura moderna. La palabra "dinosaurio" se utiliza coloquialmente para referirse a algo muy viejo o muy grande. El interés del público por estos reptiles llevó a su entrada inevitable en la cultura popular. Comenzando con una mención de pasada del megalosaurio en el primer párrafo de Casa desolada de Charles Dickens en 1852,[246] los dinosaurios han aparecido en una gran variedad de obras de ficción. El libro de 1864 de Julio Verne con Viaje al centro de la Tierra, el libro de 1912 de Arthur Conan Doyle con El mundo perdido, la icónica película de 1933 King Kong, la destacada cinta de 1953 The Beast from 20,000 Fathoms, la película japonesa de 1954 Godzilla y sus múltiples secuelas, el best seller de 1990 Parque Jurásico y su versión en película, que fue durante un corto tiempo la película más taquillera de la historia, son solo algunos ejemplos prominentes de la larga tradición de los dinosaurios en la ficción. Los dinosaurios están omnipresentes en la publicidad, y muchas compañías intentan utilizar los dinosaurios para vender sus propios productos o para tachar a sus rivales como de movimientos lentos, difumados u obsoletos.[247]
Los dinosaurios también han aparecido en los dibujos animados, desde la primera adaptación animada de los mismos con Gertie the Dinosaur (1914) hasta en la célebre cinta animada de 1940 Fantasía. Destacando, además, la dinosauromanía provocada por la película animada de George Lucas y Steven Spielberg con The Land Before Time (1988). La película, que sigue las aventuras de un grupo de jóvenes dinosaurios para llegar al «Gran Valle», tuvo tanto éxito que en septiembre de 2008, veinte años después de la original, ya habían salido doce secuelas. Asimismo, la temática dinosauria volvería a ser usada en la película We're Back! A Dinosaur's Story (1993). Los dinosaurios también han hecho apariciones en distintas series televisivas, como en Los Picapiedra, Power Rangers, Extreme Dinosaurs o Dino Rey, así como en películas de animación digital como Dinosaurio (2000), Ice Age: Dawn of the Dinosaurs (2009) y The Good Dinosaur (2015). Algunos programas infantiles destacados sobre dinosaurios fueron: Denver, el último dinosaurio, Barney y sus amigos, Dinotren y Gigantosaurus. Finalmente, cabe destacar que la reducción de los costes asociados a los efectos especiales ha permitido la creación de documentales televisivos sobre los dinosaurios, como Walking with Dinosaurs, When Dinosaurs Roamed America o Prehistoric Planet, que reproducen el aspecto de los dinosaurios con una fidelidad sin precedentes.
Al comienzo del siglo XXI, las excavaciones paleontológicas las comenzaron a organizar activamente en China y Mongolia, donde fue encontrada una multitud de otras especies de dinosaurios alados que habitaban durante el período cretácico. Este período se puede llamar condicionalmente el segundo renacimiento que siguió después del primero, en los años 1970.[248] El descubrimiento que los dinosaurios eran antepasados de las aves y tenían un comportamiento más perfecto en comparación con los reptiles simples profocó de nuevo el interés público y ejerció influencia sobre los trabajos de la nueva generación de paleoartistas.[249] En 2014, se estrenó la película australiana Dinosaur Island, que llegó a ser la primera película no documental que mostraba los dinosaurios alados.[250]
La palabra dinosaurio también se usa en el idioma inglés como despectiva, para designar a la gente, las ideas o el estilo que se hicieron anticuados hace mucho. Por ejemplo, los miembros de movimiento "punk" se han mofado del estilo de los representantes del movimiento "rock progresivo", llamándolos «dinosaurios».[251]
Concepciones religiosas
[editar]Varios grupos religiosos tienen puntos de vista sobre los dinosaurios que difieren de los que tienen la inmensa mayoría de científicos, generalmente debido a los conflictos con las historias de la Creación de sus escritos sagrados. Sin embargo, la mayoría de la comunidad científica rechaza estas interpretaciones de los dinosaurios inspiradas por la religión.[252][253]
Como es un término moderno derivado del griego, la Biblia no utiliza la palabra «dinosaurio», pero algunos cristianos interpretaron la palabra hebrea tanniyn /tan·ˈnin/ como una referencia a los dinosaurios.[254] En las traducciones al español, tanniyn puede traducirse como ‘monstruo marino’ o ‘serpiente’, pero se suele traducir como ‘dragón’. Estas criaturas son mencionadas casi treinta veces en el Antiguo Testamento, y se dice que viven tanto en la tierra como en el agua. En otro momento, la Biblia describe una criatura enorme llamada Behemot (Job 40: 15-24), que «conserva la cola tiesa, como un cedro»; el Behemot es descrita como primera entre las obras de Dios e imposible de capturar (v. 24). Algunos estudiosos de la Biblia identifican el Behemot como un elefante, un hipopótamo o un toro, pero como estos animales tienen colas muy delgadas, no se las puede comparar a la medida de un cedro; los creacionistas identifican generalmente el Behemot con los dinosaurios saurópodos. Otros creacionistas se refieren al Behemot como un Brachiosaurus, pues la Biblia dice a Job que «es la obra maestra de Dios», afirmando que es el animal más grande que Dios creó.[254] Sin embargo, algunos estudiosos creen que la referencia al cedro se refiere en realidad a sus hojas en forma de aguja, que se asemejan al pelo puntiagudo presente en la cola de los elefantes, rinocerontes e hipopótamos modernos.[255] Otros críticos afirman que la palabra «cola» es un eufemismo por el pene del animal, y que debe entenderse como una descripción de su virilidad.[256][257]
El leviatán es otra criatura mencionada en el Antiguo Testamento; su descripción le atribuye una variedad de características de dinosaurio, de dragón, y de serpiente marina. Algunos estudiosos identifican el leviatán de Job 41 con el cocodrilo del Nilo, o señalan que tiene siete cabezas y que es puramente mitológico. Como en el caso del Behemot, los creacionistas han intentado en algunas ocasiones relacionar el leviatán con los dinosaurios.[258]
Muchos musulmanes también reconocen la existencia de los dinosaurios y de otros animales extintos dentro de los límites de creacionismo islámico, según el cual Alá habría creado las «criaturas de diferentes especies» (incluso los dinosaurios) habiendo colonizado con ellos toda la Tierra. Sin embargo, se extinguieron antes de que fuese creado el primer humano.[259]
Véase también
[editar] Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios.- Fósil
- Historia de la vida
- Paleontología
- Lista de géneros válidos de dinosaurios
- Dinosaurio viviente
Referencias
[editar]- ↑ Matthew G. Baron; Megan E. Williams (2018). "A re-evaluation of the enigmatic dinosauriform Caseosaurus crosbyensis from the Late Triassic of Texas, USA and its implications for early dinosaur evolution". Acta Palaeontologica Polonica. 63. doi 10.4202/app.00372.2017.
- ↑ Andrea Cau (2018). "The assembly of the avian body plan: a 160-million-year long process" (PDF). Bollettino della Società Paleontologica Italiana. 57 (1): 1–25. doi 10.4435/BSPI.2018.01.
- ↑ Ferigolo, Jorge; Langer, Max C. (1 de enero de 2007). «A Late Triassic dinosauriform from south Brazil and the origin of the ornithischian predentary bone». Historical Biology 19 (1): 23-33. ISSN 0891-2963. S2CID 85819339. doi:10.1080/08912960600845767.
- ↑ Langer, Max C.; Ferigolo, Jorge (1 de enero de 2013). «The Late Triassic dinosauromorph Sacisaurus agudoensis (Caturrita Formation; Rio Grande do Sul, Brazil): anatomy and affinities». Geological Society, London, Special Publications (en inglés) 379 (1): 353-392. Bibcode:2013GSLSP.379..353L. ISSN 0305-8719. S2CID 131414332. doi:10.1144/SP379.16.
- ↑ Cabreira, S.F.; Kellner, A.W.A.; Dias-da-Silva, S.; da Silva, L.R.; Bronzati, M.; de Almeida Marsola, J.C.; Müller, R.T.; de Souza Bittencourt, J.; Batista, B.J.; Raugust, T.; Carrilho, R.; Brodt, A.; Langer, M.C. (2016). «A Unique Late Triassic Dinosauromorph Assemblage Reveals Dinosaur Ancestral Anatomy and Diet». Current Biology 26 (22): 3090-3095. PMID 27839975. doi:10.1016/j.cub.2016.09.040.
- ↑ Müller, Rodrigo Temp; Garcia, Maurício Silva (26 de agosto de 2020). «A paraphyletic 'Silesauridae' as an alternative hypothesis for the initial radiation of ornithischian dinosaurs». Biology Letters 16 (8): 20200417. PMC 7480155. PMID 32842895. doi:10.1098/rsbl.2020.0417.
- ↑ Welbourne, Dustin. «There's no such thing as reptiles any more – and here's why». The Conversation (en inglés). Consultado el 16 de noviembre de 2017.
- ↑ Welbourne, Dustin. «There’s No Such Thing As Reptiles Any More – And Here’s Why | Richard Dawkins Foundation». www.richarddawkins.net (en inglés estadounidense). Archivado desde el original el 16 de noviembre de 2017. Consultado el 16 de noviembre de 2017.
- ↑ Nesbitt, Sterling J.; Barrett, Paul M.; Werning, Sarah; Sidor, Christian A.; Charig, Alan J. (5 de diciembre de 2012). «The oldest dinosaur? A Middle Triassic dinosauriform from Tanzania». Biology Letters (en inglés) 9 (1): 20120949. ISSN 1744-9561. PMC 3565515. PMID 23221875. doi:10.1098/rsbl.2012.0949.
- ↑ Alcobar, Oscar A.; Martinez, Ricardo N. (19 de octubre de 2010). «A new herrerasaurid (Dinosauria, Saurischia) from the Upper Triassic Ischigualasto Formation of northwestern Argentina». ZooKeys (en inglés) 63 (63): 55-81. PMC 3088398. PMID 21594020. doi:10.3897/zookeys.63.550.
- ↑ a b c Feduccia, A. (2002). «Birds are dinosaurs: simple answer to a complex problem». The Auk (en inglés) 119 (4): 1187-1201. doi:10.1642/0004-8038(2002)119[1187:BADSAT]2.0.CO;2.
- ↑ a b c Holtz, Thomas R. Jr. (2011) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2011 Appendix. (en inglés)
- ↑ a b Wang, S.C., and Dodson, P. (2006). «Estimating the Diversity of Dinosaurs». Proceedings of the National Academy of Sciences USA (en inglés) 103 (37): 13601-13605. Bibcode:2006PNAS..10313601W. PMC 1564218. PMID 16954187. doi:10.1073/pnas.0606028103.
- ↑ Amos J (17 de septiembre de 2008). «Will the real dinosaurs stand up?». BBC News. Consultado el 23 de marzo de 2011.
- ↑ a b c d Macleod, N.; Rawson, P. F.; Forey, P. L.; Banner, F. T.; Boudagher-Fadel, M. K.; Bown, P. R.; Burnett, J. A.; Chambers, P.; Culver, S.; Evans, S. E.; Jeffery, C.; Kaminski, M. A.; Lord, A. R.; Milner, A. C.; Milner, A. R.; Morris, N.; Owen, E.; Rosen, B. R.; Smith, A. B.; Taylor, P. D.; Urquhart, E.; Young, J. R. (1997). «The Cretaceous-Tertiary biotic transition». Journal of the Geological Society (en inglés) 154 (2): 265-292. doi:10.1144/gsjgs.154.2.0265.
- ↑ a b c Benton, Michael J. (2004). «Origin and relationships of Dinosauria». En Weishampel, David B.; Dodson, Peter; y Osmólska, Halszka (eds.), ed. The Dinosauria (en inglés) (2da edición). Berkeley: University of California Press. pp. 7-19. ISBN 0-520-24209-2.
- ↑ Olshevsky, G. (2000). «An annotated checklist of dinosaur species by continent». Mesozoic Meanderings (en inglés) 3: 1-157.
- ↑ a b Langer, Max C.; Martin D. Ezcurra; Jonathas S. Bittencourt; Fernando E. Novas (2010). «The origin and early evolution of dinosaurs». Biological Reviews (en inglés) 85 (1): 65-66, 82. Archivado desde el original el 29 de octubre de 2013.
- ↑ Padian, K. (2004). "Basal Avialae". In Weishampel, D.B.; Dodson, P.; Osmolska, H. (eds.). The Dinosauria (Second ed.). Berkeley: University of California Press. pp. 210–231. ISBN 0-520-24209-2 (en inglés)
- ↑ Wade, Nicholas (22 de marzo de 2017). «Shaking Up the Dinosaur Family Tree». The New York Times (New York). ISSN 0362-4331. Archivado desde el original el 7 de abril de 2018. Consultado el 30 de octubre de 2019. (requiere registro). "A version of this article appears in print on March 28, 2017, on Page D6 of the New York edition with the headline: Shaking Up the Dinosaur Family Tree."
- ↑ Baron, Matthew G.; Norman, David B.; Barrett, Paul M. (2017). «A new hypothesis of dinosaur relationships and early dinosaur evolution». Nature (London: Nature Research) 543 (7646): 501-506. Bibcode:2017Natur.543..501B. ISSN 0028-0836. PMID 28332513. S2CID 205254710. doi:10.1038/nature21700.
- ↑ «Supplementary Information» (pdf) (en inglés). Archivado desde el original el 19 de diciembre de 2019. (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
- ↑ Max C. Langer; Martín D. Ezcurra; Oliver W. M. Rauhut; Michael J. Benton; Fabien Knoll; Blair W. McPhee; Fernando E. Novas; Diego Pol et al. (2017). «Untangling the dinosaur family tree». Nature 551 (7678): E1-E3. doi:10.1038/nature24011.
- ↑ Matthew G. Baron; David B. Norman; Paul M. Barrett (2017). «Baron et al. reply». Nature 551 (7678): E4-E5. doi:10.1038/nature24012.
- ↑ Glut, Donald F. (1997). Dinosaurs: The Encyclopedia (en inglés). Jefferson, North Carolina: McFarland & Co. p. 40. ISBN 0-89950-917-7.
- ↑ a b Lambert, David; the Diagram Group (1990). The Dinosaur Data Book (en inglés). New York: Avon Books. p. 288. ISBN 0-380-75896-2.
- ↑ Morales, Michael (1997). «Nondinosaurian vertebrates of the Mesozoic». En Farlow JO, Brett-Surman MK, ed. The Complete Dinosaur (en inglés). Bloomington: Indiana University Press. pp. 607-624. ISBN 0-253-33349-0.
- ↑ a b Tennant, Jonathan P.; Chiarenza, Alfio Alessandro; Baron, Matthew (19 de febrero de 2018). «How has our knowledge of dinosaur diversity through geologic time changed through research history?». PeerJ 6: e4417. PMC 5822849. PMID 29479504. doi:10.7717/peerj.4417.
- ↑ Starrfelt, Jostein; Liow, Lee Hsiang (2016). «How many dinosaur species were there? Fossil bias and true richness estimated using a Poisson sampling model». Philosophical Transactions of the Royal Society B: Biological Sciences 371 (1691): 20150219. PMC 4810813. PMID 26977060. doi:10.1098/rstb.2015.0219.
- ↑ Russell, Dale A. (1995). «China and the lost worlds of the dinosaurian era». Historical Biology 10: 3-12. doi:10.1080/10292389509380510.
- ↑ Starrfelt, Jostein; Liow, Lee Hsiang (2016). «How many dinosaur species were there? Fossil bias and true richness estimated using a Poisson sampling model». Philosophical Transactions of the Royal Society B (London: Royal Society) 371 (1691): 20150219. ISSN 0962-8436. PMC 4810813. PMID 26977060. doi:10.1098/rstb.2015.0219.
- ↑ Black, Riley (23 de marzo de 2016). «Most Dinosaur Species Are Still Undiscovered». National Geographic News. Archivado desde el original el 6 de marzo de 2021. Consultado el 6 de junio de 2021. (requiere registro).
- ↑ Gill, F.; Donsker, D.; Rasmussen, P. (2021). «Welcome». IOC World Bird List 11.1.
- ↑ Amiot, R.; Buffetaut, E.; Lécuyer, C.; Wang, X.; Boudad, L.; Ding, Z.; Fourel, F.; Hutt, S.; Martineau, F.; Medeiros, A.; Mo, J.; Simon, L.; Suteethorn, V.; Sweetman, S.; Tong, H.; Zhang, F.; and Zhou, Z. (2010). «Oxygen isotope evidence for semi-aquatic habits among spinosaurid theropods». Geology 38 (2): 139-142. doi:10.1130/G30402.1.
- ↑ Brusatte, Stephen L. (2012). Dinosaur Paleobiology (en inglés) (1. edición). Nueva York: Wiley, J. pp. 9-20, 21. ISBN 978-0-470-65658-7.
- ↑ Nesbitt, S.J. (2011). «The early evolution of archosaurs: relationships and the origin of major clades». Bulletin of the American Museum of Natural History 352: 1-292. doi:10.1206/352.1.
- ↑ a b Holtz, Jr., T.R. (2000). «Classification and evolution of the dinosaur groups». En Paul, G.S., ed. The Scientific American Book of Dinosaurs (en inglés). St. Martin's Press. pp. 140-168. ISBN 0-312-26226-4.
- ↑ Nesbitt, Sterling J.; Irmis, Randall B.; Parker, William G. (2007). «A critical re-evaluation of the Late Triassic dinosaur taxa of North America». Journal of Systematic Palaeontology (en inglés) 5 (2): 209-243. doi:10.1017/S1477201907002040.
- ↑ Esto no se reconoció hasta después de 1909: «Dr. Holland and the Sprawling Sauropods» (en inglés). Archivado desde el original el 12 de junio de 2011. En Desmond, A. (1976). Hot Blooded Dinosaurs (en inglés). DoubleDay. ISBN 0-385-27063-1.
- ↑ Benton, M.J. (2004). Vertebrate Paleontology (en inglés). Blackwell Publishers. xii-452. ISBN 0-632-05614-2.
- ↑ Cowen, Richard (2004). «Dinosaurs». History of Life (en inglés) (4ta edición). Blackwell Publishing. pp. 151-175. ISBN 1-4051-1756-7. OCLC 53970577.
- ↑ a b Kubo, T.; Benton, Michael J. (2007). «Evolution of hindlimb posture in archosaurs: limb stresses in extinct vertebrates». Palaeontology (en inglés) 50 (6): 1519-1529. doi:10.1111/j.1475-4983.2007.00723.x.
- ↑ Dong Zhiming (1992). Dinosaurian Faunas of China (en inglés). China Ocean Press, Beijing. ISBN 3-540-52084-8.
- ↑ BBC News, ed. (6 de julio de 2007). «Dinosaur bones 'used as medicine'» (en inglés).
- ↑ Paul, 2000, chpt. 1: "A Brief History of Dinosaur Paleontology" by Michael J. Benton.
- ↑ Plot, 1677, p. [1]
- ↑ «Robert Plot». Learning more. Oxford: Oxford University Museum of Natural History. 2006. Archivado desde el original el 1 de octubre de 2006. Consultado el 14 de noviembre de 2019.
- ↑ Lhuyd, 1699, p. 67
- ↑ Delair, Justin B.; Sarjeant, William A.S. (2002). «The earliest discoveries of dinosaurs: the records re-examined». Proceedings of the Geologists' Association (Amsterdam: Elsevier on behalf of the Geologists' Association) 113 (3): 185-197. Bibcode:2002PrGA..113..185D. ISSN 0016-7878. doi:10.1016/S0016-7878(02)80022-0.
- ↑ Gunther, 1968
- ↑ Sarjeant, William A.S. (1997). «The earliest discoveries». En Farlow, James O.; y Brett-Surman, Michael K. (editores), ed. The Complete Dinosaur (en inglés). Bloomington: Indiana University Press. pp. 3-11. ISBN 0-253-33349-0.
- ↑ Buckland, William (1824). «Notice on the Megalosaurus or great Fossil Lizard of Stonesfield». Transactions of the Geological Society of London (London: Geological Society of London) 1 (2): 390-396. ISSN 2042-5295. S2CID 129920045. doi:10.1144/transgslb.1.2.390. Archivado desde el original el 21 de octubre de 2019. Consultado el 5 de noviembre de 2019.
- ↑ Mantell, Gideon A. (1825). «Notice on the Iguanodon, a newly discovered fossil reptile, from the sandstone of Tilgate forest, in Sussex». Philosophical Transactions of the Royal Society of London (London: Royal Society) 115: 179-186. Bibcode:1825RSPT..115..179M. ISSN 0261-0523. JSTOR 107739. doi:10.1098/rstl.1825.0010.
- ↑ a b Owen, R (1842). Report on British Fossil Reptiles." Part II. Report of the Eleventh Meeting of the British Association for the Advancement of Science; Held at Plymouth in July 1841 (en inglés). Londres: John Murray. pp. 60-204.
- ↑ «Liddell–Scott–Jones Lexicon of Classical Greek» (en inglés). Consultado el 5 de agosto de 2008.
- ↑ Farlow, J.O., and Brett-Surman, M.K. (1997). «Preface». En Farlow, J.O., and Brett-Surman, M.K. (eds.), ed. The Complete Dinosaur (en inglés). Indiana University Press. pp. ix-xi. ISBN 0-253-33349-0.
- ↑ Prieto-Marques, A.; Weishampel, D.B.; Horner, J.R. (2006). The dinosaur Hadrosaurus foulkii, from the Campanian of the East Coast of North America, with a reevaluation of the genus (en inglés) (Acta Palaeontologica Polonica) 51 (1): 77-98.
- ↑ Holmes T (1996). Fossil Feud: The Bone Wars of Cope and Marsh, Pioneers in Dinosaur Science (en inglés). Silver Burdett Press. ISBN 978-0382391477.
- ↑ Diaz Martinez, I. "Dinosaurios de la Patagonia MÓDULO 1: Historia de las investigaciones de dinosaurios en Argentina". Universidad Nacional de Río Negro https://rid.unrn.edu.ar/bitstream/20.500.12049/2867/2/Modulo%201.pdf
- ↑ Taylor, M.P. (2010). «Sauropod dinosaur research: a historical review». Geological Society, London, Special Publications 343 (1): 361-386. Bibcode:2010GSLSP.343..361T. S2CID 910635. doi:10.1144/SP343.22.
- ↑ Naish, D. (2009). The Great Dinosaur Discoveries. London, UK: A & C Black Publishers Ltd. pp. 89–93. ISBN 978-1-4081-1906-8.
- ↑ Arbour, V. (2018). «Results roll in from the dinosaur renaissance». Science 360 (6389): 611. Bibcode:2018Sci...360..611A. S2CID 46887409. doi:10.1126/science.aat0451.
- ↑ a b Benton, M.J. (2008). «Fossil quality and naming dinosaurs». Biology Letters 4 (6): 729-732. PMC 2614166. PMID 18796391. doi:10.1098/rsbl.2008.0402.
- ↑ a b Cashmore, D.D.; Mannion, P.D.; Upchurch, P.; Butler, R.J. (2020). «Ten more years of discovery: revisiting the quality of the sauropodomorph dinosaur fossil record». Palaeontology 63 (6): 951-978. Bibcode:2020Palgy..63..951C. S2CID 219090716. doi:10.1111/pala.12496.
- ↑ Cashmore, D.D.; Butler, R.J. (2019). «Skeletal completeness of the non-avian theropod dinosaur fossil record». Palaeontology 62 (6): 951-981. S2CID 197571209. doi:10.1111/pala.12436.
- ↑ Holtz, T.R. Jr.; Brett-Surman, M.K. (1997). «The Taxonomy and Systematics of Dinosaurs». The Complete Dinosaur. Bloomington: Indiana University Press. pp. 209-223. ISBN 978-0-253-33349-0.
- ↑ St. Fleur, Nicholas (8 de diciembre de 2016). «That Thing With Feathers Trapped in Amber? It Was a Dinosaur Tail». Trilobites. The New York Times (New York). ISSN 0362-4331. Archivado desde el original el 31 de agosto de 2017. Consultado el 8 de diciembre de 2016. (requiere registro).
- ↑ Lockley, M.G.; Wright, J.L. (2000). «Reading About Dinosaurs – An Annotated Bibliography of Books». Journal of Geoscience Education 48 (2): 167-178. Bibcode:2000JGeEd..48..167L. S2CID 151426669. doi:10.5408/1089-9995-48.2.167.
- ↑ Lloyd, G.T.; Davis, K.E.; Pisani, D.; Tarver, J.E.; Ruta, R.; Sakamoto, M.; Hone, D.W.E.; Jennings, R. et al. (2008). «Dinosaurs and the Cretaceous Terrestrial Revolution». Proceedings of the Royal Society B 275 (1650): 2483-2490. PMC 2603200. PMID 18647715. doi:10.1098/rspb.2008.0715.
- ↑ a b Schweitzer, M.H. (2011). «Soft Tissue Preservation in Terrestrial Mesozoic Vertebrates». Annual Review of Earth and Planetary Sciences 39: 187-216. Bibcode:2011AREPS..39..187S. doi:10.1146/annurev-earth-040610-133502.
- ↑ a b Osborn, H.F. (1912). «Integument of the iguanodont dinosaur Trachodon». Memoirs of the American Museum of Natural History 1: 33-54.
- ↑ Hooley, R.W. (1917). «II—On the Integument of Iguanodon bernissartensis, Boulenger, and of Morosaurus becklesii, Mantell». Geological Magazine 4 (4): 148-150. Bibcode:1917GeoM....4..148H. S2CID 129640665. doi:10.1017/s0016756800192386.
- ↑ Bell, P.R. (2014). «A review of hadrosaur skin impressions». En Eberth, D.; Evans, D., eds. The Hadrosaurs: Proceedings of the International Hadrosaur Symposium. Bloomington: Princeton University Press. pp. 572-590.
- ↑ Eliason, C.M.; Hudson, L.; Watts, T.; Garza, H.; Clarke, J.A. (2017). «Exceptional preservation and the fossil record of tetrapod integument». Proceedings of the Royal Society B 284 (1862): 1-10. PMC 5597822. PMID 28878057. doi:10.1098/rspb.2017.0556.
- ↑ Benton, M.J. (1998). «Dinosaur fossils with soft parts». Trends in Ecology & Evolution 13 (8): 303-304. PMID 21238317. doi:10.1016/s0169-5347(98)01420-7.
- ↑ Zhou, Z.-H.; Wang, Y. (2017). «Vertebrate assemblages of the Jurassic Yanliao Biota and the Early Cretaceous Jehol Biota: Comparisons and implications». Palaeoworld 26 (2): 241-252. doi:10.1016/j.palwor.2017.01.002.
- ↑ Norell, M.A.; Xu, X. (2005). «Feathered Dinosaurs». Annual Review of Earth and Planetary Sciences 33: 277-299. Bibcode:2005AREPS..33..277N. doi:10.1146/annurev.earth.33.092203.122511.
- ↑ a b Roy, A.; Pittman, M.; Saitta, E.T.; Kaye, T.G.; Xu, X. (2020). «Recent advances in amniote palaeocolour reconstruction and a framework for future research». Biological Reviews 95 (1): 22-50. PMC 7004074. PMID 31538399. doi:10.1111/brv.12552.
- ↑ Vinther, J. (2020). «Reconstructing Vertebrate Paleocolor». Annual Review of Earth and Planetary Sciences 48: 345-375. Bibcode:2020AREPS..48..345V. S2CID 219768255. doi:10.1146/annurev-earth-073019-045641.
- ↑ Zhang, F.; Kearns, S.L.; Orr, P.J.; Benton, M.J.; Zhou, Z.; Johnson, D.; Xu, X.; Wang, X. (2010). «Fossilized melanosomes and the colour of Cretaceous dinosaurs and birds». Nature 463 (7284): 1075-1078. Bibcode:2010Natur.463.1075Z. PMID 20107440. S2CID 205219587. doi:10.1038/nature08740.
- ↑ Smithwick, F.M.; Nicholls, R.; Cuthill, I.C.; Vinther, J. (2017). «Countershading and Stripes in the Theropod Dinosaur Sinosauropteryx Reveal Heterogeneous Habitats in the Early Cretaceous Jehol Biota». Current Biology 27 (21): 3337-3343.e2. Bibcode:2017CBio...27E3337S. PMID 29107548. doi:10.1016/j.cub.2017.09.032. hdl:1983/8ee95b15-5793-42ad-8e57-da6524635349.
- ↑ Vinther, J.; Nicholls, R.; Lautenschlager, S.; Pittman, M.; Kaye, T.G.; Rayfield, E.; Mayr, G.; Cuthill, I.C. (2016). «3D Camouflage in an Ornithischian Dinosaur». Current Biology 26 (18): 2456-2462. Bibcode:2016CBio...26.2456V. PMC 5049543. PMID 27641767. doi:10.1016/j.cub.2016.06.065.
- ↑ Lindgren, J.; Moyer, A.; Schweitzer, M.H.; Sjövall, P.; Uvdal, P.; Nilsson, D.E.; Heimdal, J.; Engdahl, A.; Gren, J.A.; Schultz, B.P.; Kear, B.P. (2015). «Interpreting melanin-based coloration through deep time: a critical review». Proceedings of the Royal Society B 282 (1813): 20150614. PMC 4632609. PMID 26290071. doi:10.1098/rspb.2015.0614.
- ↑ Schweitzer, M.H.; Lindgren, J.; Moyer, A.E. (2015). «Melanosomes and ancient coloration re-examined: a response to Vinther 2015 (DOI 10.1002/bies.201500018)». BioEssays 37 (11): 1174-1183. PMID 26434749. S2CID 45178498. doi:10.1002/bies.201500061.
- ↑ Zhou, Z. (2014). «The Jehol Biota, an Early Cretaceous terrestrial Lagerstätte: new discoveries and implications». National Science Review 1 (4): 543-559. doi:10.1093/nsr/nwu055.
- ↑ O'Connor, J.K.; Zhou, Z. (2019). «The evolution of the modern avian digestive system: insights from paravian fossils from the Yanliao and Jehol biotas». Palaeontology 63 (1): 13-27. S2CID 210265348. doi:10.1111/pala.12453.
- ↑ Morell, V. (1993). «Dino DNA: the Hunt and the Hype». Science 261 (5118): 160-162. Bibcode:1993Sci...261..160M. PMID 8327889. doi:10.1126/science.8327889.
- ↑ Pawlicki, R.; Korbel, A.; Kubiak, H. (1996). «Cells, Collagen Fibrils and Vessels in Dinosaur Bone». Nature 211 (5049): 655-657. PMID 5968744. S2CID 4181847. doi:10.1038/211655a0.
- ↑ Pawlicki, R.; Nowogrodzka-Zagórska, M. (1998). «Blood vessels and red blood cells preserved in dinosaur bones». Annals of Anatomy – Anatomischer Anzeiger 180 (1): 73-77. PMID 9488909. doi:10.1016/S0940-9602(98)80140-4.
- ↑ Anderson, L.A. (2023). «A chemical framework for the preservation of fossil vertebrate cells and soft tissues». Earth-Science Reviews 240: 104367. Bibcode:2023ESRv..24004367A. S2CID 257326012. doi:10.1016/j.earscirev.2023.104367.
- ↑ a b Schweitzer, M.H.; Zheng, W.; Organ, C.L.; Avci, R.; Suo, Z.; Freimark, L.M.; LeBleu, V.S.; Duncan, M.B.; van der Heiden, M.G.; Neveu, J.M.; Lane, W.S.; Cottrell, J.S.; Horner, J.R.; Cantley, L.C.; Kalluri, R.; Asara, J.M. (2009). «Biomolecular characterization and protein sequences of the Campanian hadrosaur B. canadensis». Science 324 (5927): 626-631. Bibcode:2009Sci...324..626S. PMID 19407199. S2CID 5358680. doi:10.1126/science.1165069.
- ↑ Organ, C.L.; Schweitzer, M.H.; Zheng, W.; Freimark, L.M.; Cantley, L.C.; Asara, J.M. (2008). «Molecular Phylogenetics of Mastodon and Tyrannosaurus rex». Science 320 (5875): 499. Bibcode:2008Sci...320..499O. PMID 18436782. S2CID 24971064. doi:10.1126/science.1154284.
- ↑ Schweitzer, M.H.; Zheng, W.; Cleland, T.P.; Bern, M. (2013). «Molecular analyses of dinosaur osteocytes support the presence of endogenous molecules». Bone (Amsterdam: Elsevier) 52 (1): 414-423. ISSN 8756-3282. PMID 23085295. doi:10.1016/j.bone.2012.10.010.
- ↑ Bailleul, A.M.; Zheng, W.; Horner, J.R.; Hall, B.K.; Holliday, C.M.; Schweitzer, M.H. (2020). «Evidence of proteins, chromosomes and chemical markers of DNA in exceptionally preserved dinosaur cartilage». National Science Review 7 (4): 815-822. PMC 8289162. PMID 34692099. doi:10.1093/nsr/nwz206.
- ↑ Bertazzo, S.; Maidment, S.C.R.; Kallepitis, C. et al. (2015). «Fibres and cellular structures preserved in 75-million-year-old dinosaur specimens». Nature Communications 6: 7352. Bibcode:2015NatCo...6.7352B. ISSN 2041-1723. PMC 4468865. PMID 26056764. doi:10.1038/ncomms8352.
- ↑ Kaye, T.G.; Gaugler, G.; Sawlowicz, Z. (2008). «Dinosaurian Soft Tissues Interpreted as Bacterial Biofilms». PLOS ONE 3 (7): e2808. Bibcode:2008PLoSO...3.2808K. PMC 2483347. PMID 18665236. doi:10.1371/journal.pone.0002808.
- ↑ Peterson, J.E.; Lenczewski, M.E.; Scherer, R.P. (2010). «Influence of Microbial Biofilms on the Preservation of Primary Soft Tissue in Fossil and Extant Archosaurs». PLOS ONE 5 (10): e13334. Bibcode:2010PLoSO...513334P. ISSN 1932-6203. PMC 2953520. PMID 20967227. doi:10.1371/journal.pone.0013334.
- ↑ Buckley, M.; Warwood, S.; van Dongen, B.; Kitchener, A.C.; Manning, P.L. (2017). «A fossil protein chimera; difficulties in discriminating dinosaur peptide sequences from modern cross-contamination». Proceedings of the Royal Society B 284 (1855). PMC 5454271. PMID 28566488. doi:10.1098/rspb.2017.0544.
- ↑ a b c d e f Padian K (2004). «Basal avialae». En Weishampel DB, Dodson P, Osmólska H, ed. The Dinosauria (2da edición) (en inglés). University of California Press. pp. 210-231. ISBN 0-520-24209-2.
- ↑ Tanner LH, Lucas SG & Chapman MG (2004). «Assessing the record and causes of Late Triassic extinctions» (pdf). Earth-Science Reviews 65 (1–2): 103-139. Bibcode:2004ESRv...65..103T. doi:10.1016/S0012-8252(03)00082-5. Archivado desde el original el 25 de octubre de 2007. Consultado el 22 de octubre de 2007.
- ↑ La extinción del Pérmico-Triásico: porcentaje de animales que se extinguieron. Véase comentario
- ↑ Datos del evento Pérmico-Triásico. Véase comentario
- ↑ a b Alcober, Oscar A.; Martinez, Ricardo N. (2010). «A new herrerasaurid (Dinosauria, Saurischia) from the Upper Triassic Ischigualasto Formation of northwestern Argentina» (pdf). ZooKeys (en inglés) 63 (63): 55-81. PMC 3088398. PMID 21594020. doi:10.3897/zookeys.63.550.
- ↑ a b c Sereno PC (1999). «The evolution of dinosaurs». Science (en inglés) 284 (5423): 2137-2147. PMID 10381873. doi:10.1126/science.284.5423.2137.
- ↑ Hayward, T. The First Dinosaurs. Dinosaur Cards. Orbis Publishing Ltd. D36040612. (1997).
- ↑ a b Sereno, P.C.; Forster, Catherine A.; Rogers, Raymond R.; Monetta, Alfredo M. (1993). «Primitive dinosaur skeleton from Argentina and the early evolution of Dinosauria». Nature 361 (6407): 64-66. Bibcode:1993Natur.361...64S. doi:10.1038/361064a0.
- ↑ a b Griffin, C.T.; Wynd, B.M.; Munyikwa, D.; Broderick, T.J.; Zondo, M.; Tolan, S.; Langer, M.C.; Nesbitt, S.J. et al. (2022). «Africa's oldest dinosaurs reveal early suppression of dinosaur distribution». Nature 609 (7926): 313-319. Bibcode:2022Natur.609..313G. ISSN 0028-0836. PMID 36045297. S2CID 251977824. doi:10.1038/s41586-022-05133-x.
- ↑ Desojo, J.B.; Fiorelli, L.E.; Ezcurra, M.D.; Martinelli, A.G.; Ramezani, J.; Da Rosa, A.A.S.; Belén von Baczko, M.; Jimena Trotteyn, M.; Montefeltro, F.C.; Ezpeleta, M.; Langer, M.C. (2020). «The Late Triassic Ischigualasto Formation at Cerro Las Lajas (La Rioja, Argentina): fossil tetrapods, high-resolution chronostratigraphy, and faunal correlations». Scientific Reports 10 (1): 12782. Bibcode:2020NatSR..1012782D. PMC 7391656. PMID 32728077. doi:10.1038/s41598-020-67854-1.
- ↑ a b c Novas, F.E.; Agnolin, F.L.; Ezcurra, M.D.; Müller, R.T.; Martinelli, A.; Langer, M. (2021). «Review of the fossil record of early dinosaurs from South America, and its phylogenetic implications». Journal of South American Earth Sciences 110: 103341. Bibcode:2021JSAES.11003341N. ISSN 0895-9811. doi:10.1016/j.jsames.2021.103341.
- ↑ Müller, R.T.; Garcia, M.S. (2023). «A new silesaurid from Carnian beds of Brazil flls a gap in the radiation of avian line archosaurs». Scientific Reports 13: 4981. PMC 10090097. doi:10.1038/s41598-023-32057-x.
- ↑ Nesbitt, Sterling J; Sues, Hans-Dieter (2021). «The osteology of the early-diverging dinosaur Daemonosaurus chauliodus (Archosauria: Dinosauria) from the Coelophysis Quarry (Triassic: Rhaetian) of New Mexico and its relationships to other early dinosaurs». Zoological Journal of the Linnean Society 191 (1): 150-179. doi:10.1093/zoolinnean/zlaa080.
- ↑ a b Langer, Max C.; Ramezani, Jahandar; Da Rosa, Átila A.S. (May 2018). «U-Pb age constraints on dinosaur rise from south Brazil». Gondwana Research (Amsterdam: Elsevier) 57: 133-140. Bibcode:2018GondR..57..133L. ISSN 1342-937X. doi:10.1016/j.gr.2018.01.005.
- ↑ Novas, F.E.; Ezcurra, M.D.; Chatterjee, S.; Kutty, T.S. (2011). «New dinosaur species from the Upper Triassic Upper Maleri and Lower Dharmaram formations of central India». Earth and Environmental Science Transactions of the Royal Society of Edinburgh 101 (3–4): 333-349. Bibcode:2010EESTR.101..333N. S2CID 128620874. doi:10.1017/S1755691011020093.
- ↑ Marsicano, C.A.; Irmis, R.B.; Mancuso, A.C.; Mundil, R.; Chemale, F. (2016). «The precise temporal calibration of dinosaur origins». Proceedings of the National Academy of Sciences 113 (3): 509-513. Bibcode:2016PNAS..113..509M. PMC 4725541. PMID 26644579. doi:10.1073/pnas.1512541112.
- ↑ Nesbitt, Sterling J.; Barrett, Paul M.; Werning, Sarah et al. (2012). «The oldest dinosaur? A Middle Triassic dinosauriform from Tanzania». Biology Letters (London: Royal Society) 9 (1): 20120949. ISSN 1744-9561. PMC 3565515. PMID 23221875. doi:10.1098/rsbl.2012.0949.
- ↑ Marsicano, C.A.; Irmis, R.B.; Mancuso, A.C.; Mundil, R.; Chemale, F. (2015). «The precise temporal calibration of dinosaur origins». Proceedings of the National Academy of Sciences 113 (3): 509-513. Bibcode:2016PNAS..113..509M. ISSN 0027-8424. PMC 4725541. PMID 26644579. doi:10.1073/pnas.1512541112.
- ↑ Stephen L. Brusatte. «Superiority, Competition, and Opportunism in the Evolutionary Radiation of Dinosaurs» (en inglés).
- ↑ Justin A. Spielmann, Spencer G. Lucas and Adrian P. Hunt (2013). «The first Norian (Revueltian) rhynchosaur: Bull Canyon Formation, New Mexico, U.S.A.». New Mexico Museum of Natural History and Science Bulletin 61: 562-566. Archivado desde el original el 1 de octubre de 2015. Consultado el 18 de enero de 2015.
- ↑ Jerzy Dzik, Tomasz Sulej, and Grzegorz Niedźwiedzki (2008). «A dicynodont−theropod association in the latest Triassic of Poland». Acta Palaeontologica Polónica (en inglés) 53 (4): 733-738. doi:10.4202/app.2008.0415.
- ↑ a b c d Holtz, Thomas R., Jr.; Chapman, Ralph E.; Lamanna, Matthew C. (2004). «Mesozoic biogeography of Dinosauria». En Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.), ed. The Dinosauria (en inglés) (2da edición). Berkeley: University of California Press. pp. 627-642. ISBN 0-520-24209-2.
- ↑ a b c d e Fastovsky, David E.; Smith, Joshua B. (2004). «Dinosaur paleoecology». En Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka, ed. The Dinosauria (en inglés) (2da edición). Berkeley: University of California Press. pp. 614-626. ISBN 0-520-24209-2.
- ↑ Sereno, P.C.; Wilson, JA; Witmer, LM; Whitlock, JA; Maga, A; Ide, O; Rowe, TA; Kemp, Tom (2007). «Structural extremes in a Cretaceous dinosaur». En Kemp, Tom, ed. PLoS ONE 2 (11): e1230. Bibcode:2007PLoSO...2.1230S. PMC 2077925. PMID 18030355. doi:10.1371/journal.pone.0001230.
- ↑ Prasad, V.; Strömberg, CA; Alimohammadian, H; Sahni, A (2005). «Dinosaur coprolites and the early evolution of grasses and grazers». Science (en inglés) 310 (5751): 1170-1180. Bibcode:2005Sci...310.1177P. PMID 16293759. doi:10.1126/science.1118806.
- ↑ Archibald, J. David; Fastovsky, David E. (2004). «Dinosaur Extinction». En Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.), ed. The Dinosauria (2da edición). Berkeley: University of California Press. pp. 672-684. ISBN 0-520-24209-2.
- ↑ Lindow, B.E.K. (2011). "Bird Evolution Across the K–Pg Boundary and the Basal Neornithine Diversification." In Dyke, G. and Kaiser, G. (eds.)Living Dinosaurs: The Evolutionary History of Modern Birds, John Wiley & Sons, Ltd, Chichester, UK. doi 10.1002/9781119990475.ch14 (en inglés)
- ↑ Dal Sasso, C. y Signore, M. Exceptional soft-tissue preservation in a theropod dinosaur from Italy. Nature 292:383-387. (1998).Véase comentario
- ↑ Schweitzer, M.H., Wittmeyer, J.L. y Horner, J.R. Soft-Tissue Vessels and Cellular Preservation in Tyrannosaurus rex. Science 307:1952 - 1955. (2005). Véase comentario
- ↑ a b Alexander, R. M. (2006). "Dinosaur biomechanics". Proceedings of the Royal Society B: Biological Sciences 273 (1596): 1849–1855. doi:10.1098/rspb.2006.3532. PMC 1634776. PMID 16822743.
- ↑ Farlow, J. O.; Dodson, P.; Chinsamy, A. (1995). "Dinosaur Biology". Annual Review of Ecology and Systematics 26: 445. doi:10.1146/annurev.es.26.110195.002305.
- ↑ Weishampel, D.B., Dodson, P., Oslmolska, H. (1990). "The Dinosauria". University of California Press. pp. 733. ISBN 0-520-06727-4
- ↑ Farlow JA (1993). «On the rareness of big, fierce animals: speculations about the body sizes, population densities, and geographic ranges of predatory mammals and large, carnivorous dinosaurs». En Dodson, Peter; y Gingerich, Philip, ed. Functional Morphology and Evolution. American Journal of Science, Special Volume 293-A (en inglés). pp. 167-199.
- ↑ Peczkis, J. (1994). «Implications of body-mass estimates for dinosaurs». Journal of Vertebrate Paleontology (en inglés) 14 (4): 520-533. doi:10.1080/02724634.1995.10011575.
- ↑ «Anatomy and evolution» (en inglés). National Museum of Natural History. Consultado el 21 de noviembre de 2007.
- ↑ El tamaño de los dinosaurios varió en distintos períodos Archivado el 27 de septiembre de 2011 en Wayback Machine. Hipótesis de trabajo sobre tamaño corporal.
- ↑ «Dinosaur Giantism» (en alemán). Archivado desde el original el 19 de diciembre de 2014.
- ↑ Sander, P. Martin et al.; Christian, Andreas; Clauss, Marcus; Fechner, Regina; Gee, Carole T.; Griebeler, Eva-Maria; Gunga, Hanns-Christian; Hummel, Jürgen et al. (2011). «Biology of the sauropod dinosaurs: the evolution of gigantism». Biological Reviews 86 (1): 117-155. PMC 3045712. PMID 21251189. doi:10.1111/j.1469-185X.2010.00137.x.
- ↑ a b c Carpenter, Kenneth (2006). «Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus». En Foster, John R.; and Lucas, Spencer G. (eds.), ed. Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36 (en inglés). Albuquerque: New Mexico Museum of Natural History and Science. pp. 131-138.
- ↑ Paul, Gregory S. (2010). Princeton Field Guide to Dinosaurs. Princeton University Press. ISBN 978-0-691-13720-9.
- ↑ Colbert, Edwin Harris (1971). Men and dinosaurs: the search in field and laboratory (en inglés). Harmondsworth [Eng.]: Penguin. ISBN 0-14-021288-4.
- ↑ Mazzetta et al., G.V. (2004). «Giants and Bizarres: Body Size of Some Southern South American Cretaceous Dinosaurs.». Historical Biology (en inglés): 1-13.
- ↑ Janensch, W. (1950). The Skeleton Reconstruction of Brachiosaurus brancai. (en inglés). pp. 97-103.
- ↑ Lucas, H.; Hecket, H. (2004). «Reappraisal of Seismosaurus, a Late Jurassic Sauropod». Proceeding, Annual Meeting of the Society of Paleontology (en inglés) 36 (5): 422.
- ↑ Lovelace, David M. (2007). «Morphology of a specimen of Supersaurus (Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of diplodocid phylogeny». Arquivos do Museu Nacional (en inglés) 65 (4): 527-544.
- ↑ dal Sasso C, Maganuco S, Buffetaut E, Mendez MA (2006). «New information on the skull of the enigmatic theropod Spinosaurus, with remarks on its sizes and affinities» (pdf). Journal of Vertebrate Paleontology (en inglés) 25 (4): 888-896. doi:10.1671/0272-4634(2005)025[0888:NIOTSO]2.0.CO;2. Archivado desde el original el 29 de abril de 2011. Consultado el 5 de mayo de 2011.
- ↑ Therrien, F.; Henderson, D.M. (2007). «My theropod is bigger than yours ... or not: estimating body size from skull length in theropods». Journal of Vertebrate Paleontology (en inglés) 27 (1): 108-115. doi:10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2.
- ↑ a b Zhang F, Zhou Z, Xu X, Wang X, Sullivan C (2008). «A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers». Nature (en inglés) 455 (7216): 1105-1108. Bibcode:2008Natur.455.1105Z. PMID 18948955. doi:10.1038/nature07447.
- ↑ a b Xu X, Zhao Q, Norell M, Sullivan C, Hone D, Erickson G, Wang XL, Han FL, Guo Y (2008). «A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin». Chinese Science Bulletin (en inglés) 54 (3): 430-435. doi:10.1007/s11434-009-0009-6.
- ↑ Butler, R.J.; Zhao, Q. (2009). «The small-bodied ornithischian dinosaurs Micropachycephalosaurus hongtuyanensis and Wannanosaurus ciensis from the Late Cretaceous of China». Cretaceous Research (en inglés) 30 (1): 63-77. doi:10.1016/j.cretres.2008.03.002.
- ↑ Yans J, Dejax J, Pons D, Dupuis C & Taquet P (2005). «Implications paléontologiques et géodynamiques de la datation palynologique des sédiments à faciès wealdien de Bernissart (bassin de Mons, Belgique)». Comptes Rendus Palevol (en francés) 4 (1-2): 135-150. doi:10.1016/j.crpv.2004.12.003.
- ↑ Day, J.J.; Upchurch, P; Norman, DB; Gale, AS; Powell, HP (2002). «Sauropod trackways, evolution, and behavior». Science 296 (5573): 1659. PMID 12040187. doi:10.1126/science.1070167.
- ↑ Wright, Joanna L. (2005). «Steps in understanding sauropod biology». En Curry Rogers, Kristina A.; and Wilson, Jeffrey A., ed. The Sauropods: Evolution and Paleobiology (en inglés). Berkeley: University of California Press. pp. 252-284. ISBN 0-520-24623-3.
- ↑ Varricchio, D.J.; Sereno, Paul C.; Xijin, Zhao; Lin, Tan; Wilson, Jeffery A.; Lyon, Gabrielle H. (2008). «Mud-trapped herd captures evidence of distinctive dinosaur sociality» (pdf). Acta Palaeontologica Polonica (en inglés) 53 (4): 567-578. doi:10.4202/app.2008.0402. Consultado el 6 de mayo de 2011.
- ↑ Lessem, Don; Glut, Donald F. (1993). «Allosaurus». The Dinosaur Society's Dinosaur Encyclopedia (en inglés). Random House. pp. 19-20. ISBN 0-679-41770-2.
- ↑ Maxwell, W. D.; Ostrom, John (1995). «Taphonomy and paleobiological implications of Tenontosaurus–Deinonychus associations». Journal of Vertebrate Paleontology (en inglés) 15 (4): 707-712. doi:10.1080/02724634.1995.10011256.
- ↑ Roach, Brian T.; Brinkman, Daniel L. (2007). «A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs». Bulletin of the Peabody Museum of Natural History 48 (1): 103-138. doi:10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2.
- ↑ Tanke, Darren H. (1998). «Head-biting behavior in theropod dinosaurs: paleopathological evidence» (pdf). Gaia (en inglés) (15): 167-184. Archivado desde el original el 27 de febrero de 2008.
- ↑ «The Fighting Dinosaurs» (en inglés). American Museum of Natural History. Archivado desde el original el 18 de enero de 2012. Consultado el 5 de diciembre de 2007.
- ↑ a b Carpenter, K. (1998). «Evidence of predatory behavior by theropod dinosaurs». Gaia (en inglés) 15: 135-144.
- ↑ Rogers, Raymond R.; Krause, DW; Curry Rogers, K (2007). «Cannibalism in the Madagascan dinosaur Majungatholus atopus.». Nature (en inglés) 422 (6931): 515-518. PMID 12673249. doi:10.1038/nature01532.
- ↑ Schmitz, L.; Motani, R. (2011). «Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology». Science 332 (6030): 705-708. Bibcode:2011Sci...332..705S. PMID 21493820. doi:10.1126/science.1200043.
- ↑ Varricchio DJ, Martin, AJ and Katsura, Y (2007). «First trace and body fossil evidence of a burrowing, denning dinosaur». Proceedings of the Royal Society B (en inglés) 274 (1616): 1361-1368. PMC 2176205. PMID 17374596. doi:10.1098/rspb.2006.0443.
- ↑ Chiappe, L.M. and Witmer, L.M. (2002). Mesozoic Birds: Above the Heads of Dinosaurs. Berkeley: University of California Press. ISBN 0-520-20094-2
- ↑ Chatterjee, S.; Templin, R. J. (2007). «Biplane wing planform and flight performance of the feathered dinosaur Microraptor gui.» (pdf). Proceedings of the National Academy of Sciences (en inglés) 104 (5): 1576-1580. Bibcode:2007PNAS..104.1576C. PMC 1780066. PMID 17242354. doi:10.1073/pnas.0609975104. Archivado desde el original el 13 de octubre de 2008. Consultado el 3 de noviembre de 2015.
- ↑ Alexander RM (2006). «Dinosaur biomechanics». Proceedings of the Royal Society B (en inglés) 273 (1596): 1849-1855. PMC 1634776. PMID 16822743. doi:10.1098/rspb.2006.3532.
- ↑ Goriely A & McMillen T (2002). «Shape of a cracking whip». Physical Review Letters (en inglés) 88 (24): 244301. Bibcode:2002PhRvL..88x4301G. PMID 12059302. doi:10.1103/PhysRevLett.88.244301.
- ↑ Henderson, D.M. (2003). «Effects of stomach stones on the buoyancy and equilibrium of a floating crocodilian: A computational analysis». Canadian Journal of Zoology (en inglés) 81 (8): 1346-1357. doi:10.1139/z03-122.
- ↑ a b c d Senter, P. (2008). «Voices of the past: a review of Paleozoic and Mesozoic animal sounds». Historical Biology (en inglés) 20 (4): 255-287. doi:10.1080/08912960903033327.
- ↑ Hopson, James A. (1975). «The evolution of cranial display structures in hadrosaurian dinosaurs». Paleobiology (en inglés) 1 (1): 21-43.
- ↑ Diegert, Carl F. (1998). «A digital acoustic model of the lambeosaurine hadrosaur Parasaurolophus tubicen.». Journal of Vertebrate Paleontology (en inglés) 18 (3, Suppl.): 38A. doi:10.1080/02724634.1998.10011116.
- ↑ Currie, Philip J and Kevin Padian, ed. (1997). Encyclopedia of Dinosaurs. Academic Press. p. 206.
- ↑ Hansell M (2000). Bird Nests and Construction Behaviour. University of Cambridge Press ISBN 0-521-46038-7 (en inglés)
- ↑ Varricchio, David J.; Horner, John J.; Jackson, Frankie D. (2002). «Embryos and eggs for the Cretaceous theropod dinosaur Troodon formosus.». Journal of Vertebrate Paleontology (en inglés) 22 (3): 564-576. doi:10.1671/0272-4634(2002)022[0564:EAEFTC]2.0.CO;2.
- ↑ Lee, Andrew H.; Werning, S (2008). «Sexual maturity in growing dinosaurs does not fit reptilian growth models». Proceedings of the National Academy of Sciences (en inglés) 105 (2): 582-587. Bibcode:2008PNAS..105..582L. PMC 2206579. PMID 18195356. doi:10.1073/pnas.0708903105.
- ↑ Horner, J.R.; Makela, Robert (1979). «Nest of juveniles provides evidence of family structure among dinosaurs». Nature (en inglés) 282 (5736): 296-298. Bibcode:1979Natur.282..296H. doi:10.1038/282296a0.
- ↑ Chiappe, Luis M.; Jackson, Frankie; Coria, Rodolfo A.; Dingus, Lowell (2005). «Nesting titanosaurs from Auca Mahuevo and adjacent sites». En Curry Rogers, Kristina A.; and Wilson, Jeffrey A., ed. The Sauropods: Evolution and Paleobiology (en inglés). Berkeley: University of California Press. pp. 285-302. ISBN 0-520-24623-3.
- ↑ «Discovering Dinosaur Behavior: 1960–present view» (en inglés). Encyclopædia Britannica. Archivado desde el original el 13 de diciembre de 2013. Consultado el 5 de mayo de 2011.
- ↑ Hopp, Thomas P.; Mark J. Orsen (2004). «11: Dinosaur Brooding Behavior and the Origin of Flight Feathers». En Philip J. Currie, Eva B. Koppelhus, Martin A. Shugar, Joanna L. Wright, James O. Farlow, ed. Feathered Dragons: Studies on the Transition from Dinosaurs to Birds (en inglés). Bloomington: Indiana University Press. pp. 234-250. Consultado el 10 de diciembre de 2013.
- ↑ Reisz RR, Scott, D Sues, H-D, Evans, DC & Raath, MA (2005). «Embryos of an Early Jurassic prosauropod dinosaur and their evolutionary significance». Science (en inglés) 309 (5735): 761-764. Bibcode:2005Sci...309..761R. PMID 16051793. doi:10.1126/science.1114942.
- ↑ Clark NDL, Booth P, Booth CL, Ross DA (2004). «Dinosaur footprints from the Duntulm Formation (Bathonian, Jurassic) of the Isle of Skye» (pdf). Scottish Journal of Geology (en inglés) 40 (1): 13-21. doi:10.1144/sjg40010013. Consultado el 5 de mayo de 2011.
- ↑ Horner, J. & Gorman, J. (1988). Digging Dinosaurs: The Search that Unraveled the Mystery of Baby Dinosaurs. Workman Publishing Co.
- ↑ Peñalver, Enrique; Arillo, Antonio; Delclòs, Xavier; Peris, David; Grimaldi, David A.; Anderson, Scott R.; Nascimbene, Paul C.; Fuente, Ricardo Pérez-de la (30 de enero de 2018). «Publisher Correction: Ticks parasitised feathered dinosaurs as revealed by Cretaceous amber assemblages». Nature Communications (en inglés) 9 (1): 472. ISSN 2041-1723. PMC 5790008. PMID 29382823. doi:10.1038/s41467-018-02913-w. Consultado el 19 de abril de 2023.
- ↑ Peñalver, Enrique; Peris, David; Álvarez-Parra, Sergio; Grimaldi, David A.; Arillo, Antonio; Chiappe, Luis; Delclòs, Xavier; Alcalá, Luis et al. (25 de abril de 2023). «Symbiosis between Cretaceous dinosaurs and feather-feeding beetles». Proceedings of the National Academy of Sciences (en inglés) 120 (17): e2217872120. ISSN 0027-8424. doi:10.1073/pnas.2217872120. Consultado el 19 de abril de 2023.
- ↑ Chinsamy A, Hillenius WJ (2004). «Physiology of nonavian dinosaurs». En Weishampel DB, Dodson P, Osmólska H, ed. The Dinosauria (en inglés) (2.ª edición). University of California Press. pp. 643-659. ISBN 0-520-24209-2.
- ↑ Pontzer, H. .; Allen, V. .; Hutchinson, J. .; Farke, A. A. (2009). Farke, Andrew Allen, ed. "Biomechanics of running indicates endothermy in bipedal dinosaurs". PLoS ONE 4 (11): e7783. doi:10.1371/journal.pone.0007783. PMC 2772121. PMID 19911059.
- ↑ «Hot-Blooded or Cold-Blooded??» (en inglés). University of California Museum of Paleontology. Consultado el Febrero de 2012.
- ↑ Parsons, Keith M. (2001). Drawing out Leviathan: Dinosaurs and the science wars (en inglés). Bloomington: Indiana University Press. pp. 22-48. ISBN 0-253-33937-5.
- ↑ Sereno PC, Martinez RN, Wilson JA, Varricchio DJ, Alcober OA, et al. (Septiembre de 2008). «Evidence for Avian Intrathoracic Air Sacs in a New Predatory Dinosaur from Argentina». En Kemp, Tom, ed. PLoS ONE (en inglés) 3 (9): e3303doi=10.1371/journal.pone.0003303. Bibcode:2008PLoSO...3.3303S. PMC 2553519. PMID 18825273. doi:10.1371/journal.pone.0003303. Consultado el 39 de setiembre de 2008.
- ↑ Reid, R.E.H. (1997). «Dinosaurian Physiology: the Case for "Intermediate" Dinosaurs». En Farlow, J.O., and Brett-Surman, M.K., ed. The Complete Dinosaur. Bloomington: Indiana University Press. pp. 449-473. ISBN 0-253-33349-0.
- ↑ Ehrlich, Paul R.; David S. Dobkin; Darryl Wheye (1988). «Drinking». Birds of Stanford (en inglés). Stanford University. Consultado el 13 de diciembre de 2007.
- ↑ Tsahar, Ella; Martínez Del Rio, C; Izhaki, I; Arad, Z (2005). view=long&pmid=15767304 «Can birds be ammonotelic? Nitrogen balance and excretion in two frugivores» (Free full text). Journal of Experimental Biology 208 (6): 1025-1034. PMID 15767304. doi:10.1242/jeb.01495.
- ↑ Skadhauge, E; Erlwanger, KH; Ruziwa, SD; Dantzer, V; Elbrønd, VS; Chamunorwa, JP (2003). «Does the ostrich (Struthio camelus) coprodeum have the electrophysiological properties and microstructure of other birds?». Comparative biochemistry and physiology. Part A, Molecular & integrative physiology 134 (4): 749-755. PMID 12814783. doi:10.1016/S1095-6433(03)00006-0.
- ↑ Preest, Marion R.; Beuchat, Carol A. (Abril de 1997). «Ammonia excretion by hummingbirds». Nature 386 (6625): 561-562. Bibcode:1997Natur.386..561P. doi:10.1038/386561a0.
- ↑ Mora, J.; Martuscelli, J; Ortiz Pineda, J; Soberon, G (July 1965). «The Regulation of Urea-Biosynthesis Enzymes in Vertebrates» (pdf). Biochemical Journal 96 (1): 28-35. PMC 1206904. PMID 14343146.
- ↑ Packard, Gary C. (1966). «The Influence of Ambient Temperature and Aridity on Modes of Reproduction and Excretion of Amniote Vertebrates». The American Naturalist 100 (916): 667-682. JSTOR 2459303. doi:10.1086/282459.
- ↑ Balgooyen, Thomas G. (1 de octubre de 1971). «Pellet Regurgitation by Captive Sparrow Hawks (Falco sparverius)» (pdf). The Condor (en inglés) 73 (3): 382-385. JSTOR 1365774. doi:10.2307/1365774.
- ↑ Huxley, Thomas H. (1868). «On the animals which are most nearly intermediate between birds and reptiles». The Annals and Magazine of Natural History 4 (2): 66-75.
- ↑ Heilmann, Gerhard (1926). The Origin of Birds (en inglés). Londres: Witherby. pp. 208pp. ISBN 0-486-22784-7.
- ↑ Osborn, Henry Fairfield (1924). «Three new Theropoda, Protoceratops zone, central Mongolia» (PDF). American Museum Novitates 144: 1-12. Archivado desde el original el 27 de marzo de 2009. Consultado el 27 de septiembre de 2016.
- ↑ Ostrom, John H. (1973). «The ancestry of birds». Nature (en inglés) 242 (5393): 136. Bibcode:1973Natur.242..136O. doi:10.1038/242136a0.
- ↑ Gauthier, Jacques. (1986). «Saurischian monophyly and the origin of birds». En Padian, Kevin. (ed.), ed. The Origin of Birds and the Evolution of Flight. Memoirs of the California Academy of Sciences 8. pp. 1-55.
- ↑ Mayr, G., Pohl, B. and Peters, D.S. (2005). «A Well-Preserved Archaeopteryx Specimen with Theropod Features». Science (en inglés) 310 (5753): 1483-1486. Bibcode:2005Sci...310.1483M. PMID 16322455. doi:10.1126/science.1120331.
- ↑ María H. Schweitzer, Wenxia Zheng, Chris L Órgano, John M Asara (2009). Biomolecular Characterization and Protein Sequences of the Campanian Hadrosaur B. canadensis. Researchgate.
- ↑ John M Asara, Mary H Schweitzer, Lisa Freimark, Matthew Phillips (2007). Protein Sequences from Mastodon and Tyrannosaurus Rex Revealed by Mass Spectrometry. Researchgate.
- ↑ Martin, Larry D. (2006). «A basal archosaurian origin for birds». Acta Zoologica Sinica 50 (6): 977-990.
- ↑ a b Switek, B. (2012). «Rise of the fuzzy dinosaurs». Nature. doi:10.1038/nature.2012.10933.
- ↑ Godefroit, P., Sinitsa, S., Dhouailly, D., Bolotsky, Y., and Sizov, A. "Feather-like structures and scales in a Jurassic neornithischian dinosaur from Siberia." Program and Abstracts of the 73rd Meeting of the Society of Vertebrate Paleontology Archivado el 19 de octubre de 2013 en Wayback Machine., Octubre de 2013.
- ↑ Xu X.; Norell, M.A.; Kuang X.; Wang X.; Zhao Q.; Jia C. (2004). «Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids». Nature (en inglés) 431 (7009): 680-684. Bibcode:2004Natur.431..680X. PMID 15470426. doi:10.1038/nature02855.
- ↑ Göhlich, U.B.; Chiappe, LM (2006). «A new carnivorous dinosaur from the Late Jurassic Solnhofen archipelago». Nature (en inglés) 440 (7082): 329-332. Bibcode:2006Natur.440..329G. PMID 16541071. doi:10.1038/nature04579.
- ↑ Kellner, A.W.A., Wang, X., Tischlinger, H., Campos, D., Hone, D.W.E. and Meng, X. (2009). "The soft tissue of Jeholopterus (Pterosauria, Anurognathidae, Batrachognathinae) and the structure of the pterosaur wing membrane." Proceedings of the Royal Society B, publicado online antes de imprimirse 5 de Agosto de 2009, doi 10.1098/rspb.2009.0846
- ↑ Alibardi, L., Knapp, L.W., Sawyer, R.H. (2006). "Beta-keratin localization in developing alligator scales and feathers in relation to the development and evolution of feathers." US National Library of Medicine, Junio de 2006
- ↑ Wellnhofer, P (1988). «Ein neuer Exemplar von Archaeopteryx». Archaeopteryx 6: 1-30.
- ↑ Schweitzer, M.H.; Watt, J.A.; Avci, R.; Knapp, L.; Chiappe, L.; Norell, M.; Marshall, M. (1999). «Beta-keratin specific immunological reactivity in feather-like structures of the Cretaceous Alvarezsaurid,Shuvuuia deserti». Journal of Experimental Zoology (en inglés) 285 (2): 146-157. PMID 10440726. doi:10.1002/(SICI)1097-010X(19990815)285:2<146::AID-JEZ7>3.0.CO;2-A.
- ↑ Lingham-Soliar, T. (2003). «The dinosaurian origin of feathers: perspectives from dolphin (Cetacea) collagen fibers». Naturwissenschaften (en inglés) 90 (12): 563-567. Bibcode:2003NW.....90..563L. PMID 14676953. doi:10.1007/s00114-003-0483-7.
- ↑ a b Feduccia, A.; Lingham-Soliar, T; Hinchliffe, JR (2005). «Do feathered dinosaurs exist? Testing the hypothesis on neontological and paleontological evidence». Journal of Morphology (en inglés) 266 (2): 125-166. PMID 16217748. doi:10.1002/jmor.10382.
- ↑ Lingham-Soliar, T.; Feduccia, A; Wang, X (2007). «A new Chinese specimen indicates that 'protofeathers' in the Early Cretaceous theropod dinosaur Sinosauropteryx are degraded collagen fibres». Proceedings of the Royal Society B (en inglés) 274 (1620): 1823-1829. PMC 2270928. PMID 17521978. doi:10.1098/rspb.2007.0352.
- ↑ Prum, Richard O. (Abril de 2003). «Are Current Critiques Of The Theropod Origin Of Birds Science? Rebuttal To Feduccia 2002». The Auk (en inglés) 120 (2): 550-561. JSTOR 4090212. doi:10.1642/0004-8038(2003)120[0550:ACCOTT]2.0.CO;2.
- ↑ Archaeopteryx: a missing link. Berkeley: University of California. Museum of Paleontology.
- ↑ O'Connor PM & Claessens LPAM (2005). «Basic avian pulmonary design and flow-through ventilation in non-avian theropod dinosaurs». Nature (en inglés) 436: 253-256. doi:10.1038/nature03716.
- ↑ Sereno, P.C.; Martinez, R.N.; Wilson, J.A.; Varricchio, D.J.; Alcober, O.A. i Larsson, H.C.E. (Setiembre de 2008). «Evidence for Avian Intrathoracic Air Sacs in a New Predatory Dinosaur from Argentina». PLoS ONE (en inglés) 3 (9). doi:10.1371/journal.pone.0003303. Consultado el 27 de octubre de 2008.
- ↑ Meat-Eating Dinosaur from Argentina Had Bird-Like Breathing System Newswise, Consultado el 29 setiembre 2008. (en inglés)
- ↑ Xu, X.; Norell, M.A. (2004). «A new troodontid dinosaur from China with avian-like sleeping posture». Nature (en inglés) 431 (7010): 838-841. Bibcode:2004Natur.431..838X. PMID 15483610. doi:10.1038/nature02898.
- ↑ Norell M.A.; Clark J.M.; Chiappe L.M.; Dashzeveg D. (1995). «A nesting dinosaur». Nature (en inglés) 378 (6559): 774-776. Bibcode:1995Natur.378..774N. doi:10.1038/378774a0.
- ↑ Varricchio, D. J.; Moore, J. R.; Erickson, G. M.; Norell, M. A.; Jackson, F. D.; Borkowski, J. J. (2008). «Avian Paternal Care Had Dinosaur Origin». Science (en inglés) 322 (5909): 1826-1828. Bibcode:2008Sci...322.1826V. PMID 19095938. doi:10.1126/science.1163245.
- ↑ Wings O (2007). «A review of gastrolith function with implications for fossil vertebrates and a revised classification» (PDF). Palaeontologica Polonica (en inglés) 52 (1): 1-16. Consultado el 5 de mayo de 2011.
- ↑ Dingus, L. and Rowe, T. (1998). The Mistaken Extinction – Dinosaur Evolution and the Origin of Birds. Nueva York: W. H. Freeman.
- ↑ Longrich, NR; Bhullar, BA; Gauthier, JA (2012). «Mass extinction of lizards and snakes at the Cretaceous–Paleogene boundary». Proc. Natl. Acad. Sci. U.S.A. (en inglés) 109 (52): 396-401. Bibcode:2012PNAS..10921396L. PMC 3535637. PMID 23236177. doi:10.1073/pnas.1211526110.
- ↑ Keller, G.; Adatte, T.; Gardin, S.; Bartolini, A.; Bajpai, S. (2008). «Main Deccan volcanism phase ends near the K–T boundary: Evidence from the Krishna–Godavari Basin, SE India». Earth and Planetary Science Letters (en inglés) 268 (3-4): 293-311. Bibcode:2008E&PSL.268..293K. doi:10.1016/j.epsl.2008.01.015.
- ↑ Mullen, L. (2004). "Multiple Impacts". Astrobiology Magazine.
- ↑ MacLeod, N.; Rawson, P. F.; Forey, P. L.; Banner, F. T.; Boudagher-Fadel, M. K.; Bown, P. R.; Burnett, J. A.; Chambers, P.; Culver, S.; Evans, S. E.; Jeffery, C.; Kaminski, M. A.; Lord, A. R.; Milner, A. C.; Milner, A. R.; Morris, N.; Owen, E.; Rosen, B. R.; Smith, A. B.; Taylor, P. D.; Urquhart, E.; Young, J. R. (1997). «The Cretaceous-Tertiary biotic transition». Journal of the Geological Society (en inglés) 154 (2): 265-292. doi:10.1144/gsjgs.154.2.0265.
- ↑ Miller KG, Kominz MA, Browning JV, Wright JD, Mountain GS, Katz ME, Sugarman PJ, Cramer BS, Christie-Blick N, Pekar SF (2005). «The Phanerozoic record of global sea-level change». Science (en inglés) 310 (5752): 1293-1298. PMID 16311326. doi:10.1126/science.1116412.
- ↑ McArthura JM, Janssenb NMM, Rebouletc S, Lengd MJ, Thirlwalle MF & van de Shootbruggef B (2007). «Palaeotemperatures, polar ice-volume, and isotope stratigraphy (Mg/Ca, δ18O, δ13C, 87Sr/86Sr): The Early Cretaceous (Berriasian, Valanginian, Hauterivian)». Palaeogeography, Palaeoclimatology, Palaeoecology (en inglés) 248 (3-4): 391-430. doi:10.1016/j.palaeo.2006.12.015.
- ↑ Alvarez, LW, Alvarez, W, Asaro, F, and Michel, HV (1980). «Extraterrestrial cause for the Cretaceous–Tertiary extinction». Science (en inglés) 208 (4448): 1095-1108. PMID 17783054. doi:10.1126/science.208.4448.1095.
- ↑ Hildebrand, Alan R.; Penfield, Glen T.; Kring, David A.; Pilkington, Mark; Zanoguera, Antonio Camargo; Jacobsen, Stein B.; Boynton, William V. (Setiembre de 1991). «Chicxulub Crater; a possible Cretaceous/Tertiary boundary impact crater on the Yucatan Peninsula, Mexico». Geology (en inglés) 19 (9): 867-871. doi:10.1130/0091-7613(1991)019<0867:CCAPCT>2.3.CO;2.
- ↑ Robertson, D.S.; etal (30 de setiembre de 2003). «Survival in the first hours of the Cenozoic». Geological Society of America Bulletin (en inglés) 116 (5/6): 760-768. Bibcode:2004GSAB..116..760R. doi:10.1130/B25402.1. Archivado desde el original el 22 de octubre de 2015. Consultado el 15 de junio de 2011.
- ↑ a b Hofman, C, Féraud, G & Courtillot, V (2000). «40Ar/39Ar dating of mineral separates and whole rocks from the Western Ghats lava pile: further constraints on duration and age of the Deccan traps». Earth and Planetary Science Letters 180: 13-27. doi:10.1016/S0012-821X(00)00159-X.
- ↑ a b c Duncan, RA & Pyle, DG (1988). «Rapid eruption of the Deccan flood basalts at the Cretaceous/Tertiary boundary». Nature (en inglés) 333: 841-843. doi:10.1038/333841a0.
- ↑ Alvarez, W (1997). T. rex and the Crater of Doom (en inglés). Princeton University Press. pp. 130-146. ISBN 978-0691016306.
- ↑ Graeme T. Lloyd, Katie E. Davis, Davide Pisani, James E. Tarver, Marcello Ruta, Manabu Sakamoto, David W.E. Hone, Rachel Jennings, Michael J. Benton (noviembre de 2008). «Dinosaurs and the Cretaceous Terrestrial Revolution». Proceedings of the Royal Society (en inglés). vol. 275 (n. 1650): p. 2483-2490. Consultado el 3 de diciembre de 2016.
- ↑ Lloyd, G.T., Davis, K.E., Pisani, D., et al (22 de julio de 2008). «Dinosaurs and the Cretaceous Terrestrial Revolution». Proceedings of the Royal Society: Biology (en inglés). doi:10.1098/rspb.2008.0715. Consultado el 28 de julio de 2008.
- ↑ Fassett, J. E.; Lucas, S. G.; Zielinski, R. A. y Budahn, J. R. (2001). «Compelling new evidence for Paleocene dinosaurs in the Ojo Alamo Sandstone, San Juan Basin, New Mexico and Colorado, USA». Contribución n.º 1053. Catastrophic events and mass extinctions: Impacts and Beyond. Vienna, July 9-12, 2000 (en inglés) (Lunar and Planetary Institute): 45-46. Consultado el 18 de mayo de 2007.
- ↑ Sloan, R. E., Rigby, K,. Van Valen, L. M., Gabriel, Diane (1986). «Gradual dinosaur extinction and simultaneous ungulate radiation in the Hell Creek Formation». Science (en inglés) 232 (4750): 629-633. PMID 17781415. doi:10.1126/science.232.4750.629. Consultado el 18 de mayo de 2007.
- ↑ a b Fastovsky, David E.; and Sheehan, Peter M. (2005). «Reply to comment on "The Extinction of the dinosaurs in North America"» (pdf). GSA Today (en inglés) 15: 11. doi:10.1130/1052-5173(2005)015[11b:RTEOTD]2.0.CO;2. Archivado desde el original el 2 de junio de 2018. Consultado el 3 de diciembre de 2016.
- ↑ Sullivan, R. M. (2003). «No Paleocene dinosaurs in the San Juan Basin, New Mexico». Geological Society of America Abstracts with Programs (en inglés) 35 (5): 15. Archivado desde el original el 8 de abril de 2011. Consultado el 2 de julio de 2007.
- ↑ Torrens, H.S. (1993). «The dinosaurs and dinomania over 150 years». Modern Geology (en inglés) 18 (2): 257-286.
- ↑ Breithaupt, Brent H. (1997). First golden period in the USA." In: Currie, Philip J. & Padian, Kevin (Eds.). The Encyclopedia of Dinosaurs (en inglés). University of California Press. pp. 347-350. ISBN 978-0122268106.
- ↑ Londres. Michaelmas term lately over, and the Lord Chancellor sitting in Lincoln's Inn Hall. Implacable November weather. As much mud in the streets, as if the waters had but newly retired from the face of the earth, and it would not be wonderful to meet a Megalosaurus, forty feet long or so, waddling like an elephantine lizard up Holborne Hill. From page 1 of Dickens, Charles J.H. (1852). Bleak House. Londres: Bradbury & Evans.
- ↑ Glut, DF, & Brett-Surman, MK (1997). Dinosaurs and the media. En: Farlow, James O. & Brett-Surman, Michael K. (Eds.). The Complete Dinosaur (en inglés). Indiana University Press. pp. 675-697. ISBN 978-0253213136.
- ↑ «Artistic Depictions of Dinosaurs Have Undergone Two Revolutions». blogs.scientificamerican.com (en inglés). Consultado el 2 de julio de 2021.
- ↑ «“Dinosaur Renaissance”-Revolution in Dinosaur Research». onlydinosaurs.com (en inglés). Consultado el 2 de julio de 2021.
- ↑ «'Dinosaur Island' Will Feature A Feathered T. Rex». inquisitr.com (en inglés). Consultado el 2 de julio de 2021.
- ↑ «dinosaur». dictionary.com (en inglés). Consultado el 2 de julio de 2021.
- ↑ Kitcher, Philip (1983). Abusing Science: The Case Against Creationism (en inglés). MIT Press. pp. 213. ISBN 978-0-262-61037-7. Archivado desde el original el 23 de junio de 2008.
- ↑ Dawkins, Richard (1996). The Blind Watchmaker: Why the Evidence of Evolution Reveals a Universe Without Design (en inglés). W.W. Norton. p. 400. ISBN 978-0393315707.
- ↑ a b «Dinosaurs and the Bible» (en inglés). Clarifying Christianity'. 2005. Consultado el 14 de marzo de 2007.
- ↑ Bright, Michael (2006). Beasts of the Field: The Revealing Natural History of Animals in the Bible (en inglés). pp. p346. 1861058314.
- ↑ Pennock, Robert T. (2000). Tower of Babel: the evidence against the new creationism (en inglés). Cambridge, Mass: MIT Press. ISBN 0-262-66165-9.
- ↑ «Index to Creationist Claims» (en inglés). Consultado el 13 de setiembre de 2007.
- ↑ the TalkOrigins Archive (ed.). «Index to Creationist Claims» (en inglés). Consultado el 14 de marzo de 2007.
- ↑ «Why Did Allah Destroy the Dinosaurs?». aboutislam.net (en inglés). Consultado el 2 de julio de 2021.
Bibliografía
[editar]En español
[editar]- John Malam & Steve Parker. 2002.
- Andrés, J.A. y Pérez-Lorente, F. (2005): «La declaración de los yacimientos de icnitas de dinosaurio de la Península Ibérica como patrimonio mundial (IDPI)». XXI Jornadas de la Sociedad Española de Paleontología. Gestión e Investigación de la Paleontología en el XXI. Sevilla 4-8 de octubre de 2005.
- Bravo, A.M., Vila, B., Galobart, A. y Oms, O. (2005): «Restos de huevos de dinosaurio en el Cretácico superior del sinclinal de Vallcebre (Berguedà, provincia de Barcelona)». Revista Española de Paleontología, Número extraordinario, 10: 49-57.
- Le Loeuff, J. y Martínez, A. (1997): «Afloramiento de icnitas de Titanosauridae en la zona de Fumanya (Maastrichtiense, Pirineo oriental): estudio preliminar». Geogaceta, 21: 151-153.
- Sanz, J.L. (1999): Los dinosaurios voladores. Historia evolutiva de las aves primitivas. Ediciones Libertarias/Prodhufi, S.A. Mundo Vivo. 239 pp. ISBN 84-7954-493-7.
- Sanz, J.L. (1999): Mitología de los dinosaurios. Editorial Taurus. Pensamiento. 206 pp. ISBN 84-306-0348-4.
- Sanz, J.L. (2007): Cazadores de dragones. Historia del descubrimiento e investigación de los dinosaurios. Editorial Ariel. 420 pp. ISBN 978-84-344-5316-6.
- Sanz, J.L. y Buscalioni, A.D. (Coords.) (1992): Los dinosaurios y su entorno biótico. Ayuntamiento de Cuenca, Instituto «Juan de Valdés». Actas académicas, 4. 397 pp. ISBN 84-86788-14-5.
- Sanz, J.L., Buscalioni, A.D., Moratalla, J.J., Francés, V. y Antón, M. (1990): Los reptiles mesozoicos del registro español. C.S.I.C. Museo Nacional de Ciencias Naturales. Monografías, 2. 79 pp. ISBN 84-00-07036-4 [en los ejemplares figura el ISBN 84-7476-133-6].
En inglés
[editar]- Padian, K., y Currie, Ph.J. (1997): Encyclopedia of Dinosaurs. Academic Press. ISBN 0-12-226810-5.
- Paul, G.S. (2000): The Scientific American Book of Dinosaurs. St. Martin's Press. ISBN 0-312-26226-4.
- Paul, G.S. (2002): Dinosaurs of the Air: The Evolution and Loss of flight in Dinosaurs and Birds. The Johns Hopkins University Press. Baltimore. ISBN 0-8018-6763-0.
- Weishampel, D.B. (2004): The Dinosauria. University of California Press. California. [2.ª edición] ISBN 0-520-24209-2.
Enlaces externos
[editar] Wikimedia Commons alberga una categoría multimedia sobre Dinosauria.
Wikimedia Commons alberga una categoría multimedia sobre Dinosauria. Wikispecies tiene un artículo sobre Dinosauria.
Wikispecies tiene un artículo sobre Dinosauria.
- General
- Imágenes
- The Science and Art of Gregory S. Paul Influencia del arte y pinturas de la anatomía paleontológica (en inglés)
- Skeletal Drawing Restauraciones profesionales de numerosos dinosaurios, y los debates sobre la anatomía de dinosaurios (en inglés).
- Video
- BBC Nature: Vea dinosaurios cobran vida y encuentre interpretaciones de los expertos con videos de programas de la BBC, incluyendo Caminando entre dinosaurios (en inglés).
- BBC Explainer - Dinosaurios - una historia completa en 4 minutos, animación (en inglés).
- Popular
- Dinosaurios y otras criaturas extintas: del Museo de Historia Natural, un directorio de dinosaurios bien ilustrado (en inglés).
- Dinosaurnews (www.dinosaurnews.org) Titulares relacionados con dinosaurios de todo el mundo. Noticias recientes sobre dinosaurios, incluyendo hallazgos y descubrimientos, y muchos enlaces (en inglés).
- Dinosauria Desde el Museo de Paleontología de la Universidad de Berkeley, información detallada —desplácese hacia abajo para ir al menú— (en inglés).
- LiveScience.com Todo sobre dinosaurios, con artículos destacados vigentes (en inglés).
- Zoom Dinosaurs (www.enchantedlearning.com) Desde Enchanted Learning. Sitio infantil, páginas de info y estadísticas, teorías, historia (en inglés).
- Dinosaur genus list contiene tablas de datos en casi todos los género de dinosaurios mesozoicos publicados a partir de enero de 2011 (en inglés).
- Dinosaurios: (www.dinosaurios.wiki) Wiki informativa dedicada a los dinosaurios.
- Esta obra contiene una traducción total y parcial derivada de «Dinosaur» de Wikipedia en inglés, concretamente de esta versión, publicada por sus editores bajo la Licencia de documentación libre de GNU y la Licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional.
- Esta obra contiene una traducción total y parcial derivada de «Dinosaure» de Wikipedia en catalán, concretamente de esta versión, publicada por sus editores bajo la Licencia de documentación libre de GNU y la Licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional.
| Control de autoridades |
|
|---|
 Datos: Q430
Datos: Q430- Multimedia: Dinosauria / Q430
- Especies: Dinosauria